Introduction
S100 was first quarantined from brain tissue and first identified by B.W. Moore in 1965. It is called so, because of its solubility in 100% saturated solution of ammonium sulphate at neutral pH [1]. S100 was thought to be of a precise neural cell origin, but it is expressed in many cells [2]. S100 protein role in calcium binding was initially identified in the brain of several mammals [2,3]. Till date 24 different types of S100 proteins have been identified [4]. S100 protein was recognized in nearly all nerve sheath tumors and also expressed in controversial histogenesis tumors. Today altered levels of S100 proteins has been found to be associated with cardiomyopathies, neurogenerative disorders, inflammatory disorders, cancer and has also been reported in oral diseases such as oral melanomas, salivary microtubules in centrioles and cilia and gland tumours, granular cell myoblastoma and oral lichen planus. S100 has a broad range of intracellular and extra cellular functions [5].
Recent studies have shown the relationship of S100 proteins with different types of cancer, serving in tumorigenesis and cancer progression. The association between S100 proteins and cancer can also be elucidated by several explanations: firstly, most of S100 genes are huddled on human chromosome 1q21, a region prone to genomic rearrangements, supporting that S100 proteins may be implicated in tumor progression. Secondly, several tumors show varied S100 protein expression. Finally, a number of S100 proteins are involved in cancer by interacting with specific target proteins such as NF-κB, p53, and β-catenin [5]. Thus in this review we update the current nomenclature and chromosomal location of S100, structure, functions, types/members of S100 protein, distribution in normal tissues and the role of S100 proteins in tumorigenesis, cancer metastasis, tumor microenvironment and expression of S100 in normal and pathological conditions.
Structure of S100: S100 proteins are homologous intracellular calcium-binding type with low molecular weight (9-13 kDa) tissue and cell specific expressions, which are characterized by EF- hand motifs unique to vertebrates, with an ability to form homodimers, heterodimers and oligomeric assemblies [6]. The monomers (10-12 kDa) are either alpha or beta subunits or are each composed to two "hands" [2,7]. The dimer of two alpha subunits is termed Sl00A, that of two beta subunits SI00B, and the heterodimer of one alpha and one beta subunits S100A. The specificity of each monomer is probably determined in the C-terminal region and the "hinge" region between each EF-hand. The primary amino acid sequence of Sl00 protein was elucidated by Isobe et al., and the structure of the Sl00 gene is gradually being unraveled [2].
Types of S100: Till date 25 types of S100 proteins have been recognized [4, 8], among these 22 (S100A1–S100A18, trichohylin, filaggrin and repetin) are clustered at chromosome locus 1q21, while other S100 proteins are found at chromosome loci 4p16 (S100P), 5q14 (S100Z), 21q22 (S100B) and Xp22 (S100G) [8].
Function of S100: Even though S100 is evenly distributed in cytoplasm and in some cell organelles, role of S100 is not understood fully, but its function is probably attributed to calcium binding properties. S100 is also a constituent of microtubules in centrioles and cilia and has the ability to control microtubule assembly and disassembly in the presence of calcium and zinc invitro, thus associated with formation of microtubules by complexing with proteins. S100 also controls cell cycle by its presence in nucleoplasm [1,7].
S100 is multifunctional protein expressed in diverse spectrum of tissues, having intracellular and extracellular roles such as-
Regulation of cell proliferation and regulation [9],
Cell division [2,6],
Ca2+ homeostasis [9],
Structural organization of membranes [1],
Dynamics of cytoskeleton constituents[1],
Cell growth, motility and survival [1,8],
Enzyme activation [1],
Protection from oxidative cell damage [1],
Protein phosphorylation and secretion [1,8].
In divergence to intracellular function, extracellular S100 proteins act like cytokines by binding to cell surface receptors such as the Receptor for Advanced Glycation End products (RAGE) and Toll-Like Receptors (TLRs).
This diversity of functions appears due to [1]:
Wide diversification of the different members (24 types in human).
Different metal ion-binding properties of the individual S100 proteins.
Spatial distribution in specific intracellular compartments or extracellular space.
Their ability to form non-covalent homo and hetero-dimers, allowing for dynamic exchange of the S100 subunits.
Though intercellular function have been studied extensively, but during cell damage or any stimuli, S100 proteins can be released into extracellular space and thereby promoting various paracrine and autocrine functions such neuronal survival and extension (S100B), apoptosis (S100A4 and S100A6), inflammation (S100B, S100A8/A9, S100A11 and S100A12), autoimmunity (S100A8/A9), chemotaxis (S100A8/A9) and cell proliferation and survival (S100P, S100A7). S100B, S100P, S100A4, S100A6, S100A8/A9, S100A11 and S100A12 are known to act via interaction with cell surface receptors, primarily the Receptor for Advanced Glycation End products (RAGE), while S100A8/A9 binds to Toll-Like Receptors (TLRs). Multimeric forms of S100 proteins appear to be necessary for the extracellular functions of S100 proteins, thus elevated S100 protein levels are associated with chronic inflammation, neurodegeneration, cardiomyopathies, atherosclerosis and cancer [Table/Fig-1] [9,10].
Distribution of Sl00 protein, and subunits where known, in normal structures and cells found in and around the head and neck [9,10].
| S100Proteins | Normal Expression | Tumors in which expression is up-regulated | Tumors in which expression is down-regulated | Known interactions / Mechanism of Action / Clinical Implication |
|---|
| S100A1 | Heart Less in skeletal muscle
| Renal oncocytomas, clear cell and papillary renal cell carcinomas Endometrium subtype of ovarian and endometrial carcinomas
| ____ | A marker for poor prognosis ovarian and endometrial cancers. A specific and sensitive marker in distinguishing nephrogenic adenoma from prostatic adenocarcinoma. Known RAGE ligand.
|
| S100A2 | Many organs Increased expression in lungs and kidneys
| Non-small cell carcinoma of lung Pancreatic carcinoma Colon carcinoma Gastric carcinoma Thyroid, papillary and anaplastic carcinoma
| Larynx carcinoma Oral squamous cell carcinoma Prostate, bladder carcinoma
| A poor prognostic marker for NSCLC and pancreatic cancer A predictor of good prognosis and survival in OSCC, ESCC and LSCC. An independent prognostic marker for recurrence in oral cancer and CRC. A predictor of response pancreatectomy and a metastatic marker for pancreatic cancer. Promotes p53 transcriptional activity and reduces expression of COX-2.
|
| S100A3 | Hair cuticle
| ____ | Invasive breast tumor
| Not known |
| S100A4 also called Metastatin | Nervous system Expression is low in normal tissues
| Breast cancer Colorectal carcinoma Gastric carcinoma Prostate carcinoma Non-small cell carcinoma of lung Ovarian carcinoma Pancreatic carcinoma Melanoma carcinoma
| Oral
| Promotes tumor migration, invasion and angiogenesis. Regulates matrix metalloproteinases and interacts with p53 and inhibits p53 phosphorylation. Biomarker for metastasis in multiple cancer
|
| S100A5 | Restricted regions of the adult brain.
| Brain tumors
| ____ | ____ |
| S100A6 | Epithelial cells Fibroblasts
| Colorectal carcinoma Pancreatic carcinoma Gastric carcinoma Hepatocellular carcinoma Lung carcinoma Melanoma
| Prostate cancer and Medulloblastomas.
| Diagnostic marker or prognostic factor in pancreatic cancer, gastric cancer, prostate cancer, melanoma. Known RAGE ligand.
|
| S100A7 also called psoriasin | Epithelial cells of skin Antibacterial effect
| Breast cancer Bladder carcinoma Skin carcinoma Oral carcinoma
| Salivary gland carcinoma
| A predictor of poor prognosis in HNSCC and estrogen receptor-negative invasive breast cancer. Serum S100A7 may serve as a potentially diagnostic marker for lung cancer and ovarian cancer. Possible interaction with Jab-1.
|
| S100A8 | Expressed predominantly by myeloid cells, including granulocytes, monocytes, myeloid derived suppressor cells (MDSC) and other immature cells of myeloid lineage
| Gastric carcinoma Colon carcinoma Pancreatic carcinoma Bladder carcinoma Ovarian carcinoma Thyroid carcinoma Breast carcinoma Skin carcinoma
| Oral carcinoma Cervix carcinoma
| Interaction with RAGE and TLR4; promote tumor proliferation, and migration, accumulation of myeloid derived suppressor cells, activation of protumorigenic genes, and formation of premetastatic niches in distal organs.
|
| S100A9 | Expressed predominantly by myeloid cells, including granulocytes, monocytes, myeloid derived suppressor cells (MDSC) and other immature cells of myeloid lineage
| Breast cancer Stomach carcinoma Lung carcinoma Cervix carcinoma Prostate carcinoma Oral carcinoma
| Esophagus
| A poor prognostic marker in IDC and NSCLC; Serum S100A9 may serve as a useful marker to discriminate between prostate cancer and benign prostatic hyperplasia.
|
| S100A10-also known as p11 protein | Predominantly in lungs and kidney Brain
| Breast cancer Stomach carcinoma Gallbladder carcinoma Kidney carcinoma
| ___ | A potential molecular target for early gallbladder cancer diagnostics and therapeutic applications.
|
| S100A11 | Placenta Heart Lung Kidney
| Uterine, smooth muscle carcinoma Lymphoma, anaplastic large cell Pancreatic carcinoma
| Bladder carcinoma Esophageal, squamous cell carcinoma
| Down-regulation of S100C is associated with bladder cancer progression and poor survival.
|
| S100A13 | Thyroid gland Smooth muscle cells
| Melanoma
| ___ | An angiogenic and prognostic marker in melanoma.
|
| S100A14 | ____ | Breast cancer Ovary carcinoma Uterus carcinoma
| Oral carcinoma Kidney carcinoma Rectum carcinoma Colon carcinoma
| Promotes cell motility and invasiveness by regulating the expression and function of MMP2 in a p53-dependent manner.
|
| S100A15 also called as Koebnerisin | Epidermal basal Differentiated keratinocytes, melanocytes Langerhan’s cells
| Breast carcinoma
| ____ | Regulates tumor progression Overexpressed in inflammatory skin diseases, such as psoriasis and eczema.
|
| S100A16 | Brain (astrocytoma)
| Bladder carcinoma Lung carcinoma Thyroid carcinoma Pancreas carcinoma Ovary carcinoma
| ____ | S100A14 and S100A16 might be prognostic biomarkers and potential therapeutic targets for breast cancer
|
| S100B | Brain – highly expressed by astrocytes and schwann cells
| Brain – highly expressed by astrocytes and schwann cells
| ____ | Interacts with p53 and down-regulates p53-mediated apoptosis in melanoma. Well known RAGE ligand.
|
| S100P | Placenta GI tract Prostate Leukocyte
| Ovarian carcinoma Pancreatic carcinoma Breast cancer Gastric carcinoma Colorectal carcinoma Prostate carcinoma Lung carcinoma
| _____ | S100P is potentially useful for molecular diagnosis of NSCLCs. Elevated S100P level showed poorer overall survival in CRC and ovarian cancer patients. Activation of RAGE dependent signaling pathways.
|
NSCLC- Non-Small Cell Lung Cancer, OSCC- Oral Squamous Cell Carcinoma, ESCC- Esophagus Squamous Cell Carcinoma, LSCC- Laryngeal Squamous Cell Carcinoma, CRC- Colorectal Cancer.
Distribution of S100 in normal mineralized tissues, oral mucosa and salivary glands.
• Mineralized tissue: Most, but not all of the mesenchymal tissue in the head and neck which successively mineralizes is of neural crest cell origin. S100 distribution varies in species which may reflect degree of specialization or functional adaptation; in contrast to rodents, human odontoblasts were S100+, which was supported by Carbone et al., but in contrast few studies were unable to demonstrate S100+ in bell stage of development [7]. Chondrocytes have been found to be positive for alpha and beta subunits of S100 [1,7]. Though often reported as S100-, at least one group has found osteoblasts and osteocytes to be S100+ [1].
• Oral mucosa and salivary glands: S100 is normally expressed in normal structures of head and neck region like salivary gland, dental tissue, surface epithelium and connective tissue cells [Table/Fig-2].
Different types of S100 and their relation with different types of cancer.
| Tissue | S100α | S100β | Polyclonal |
|---|
| Salivary glands |
| Acinar cells | ± | ± | _ |
| Intercalated cells | ± | + | ± |
| Striated duct cells | - | - | _ |
| Myoepithelial cells | | ± | ± |
| Cells in and associated with surface epithelium |
| Keratinocytes | _ | _ | _ |
| Melanocytes | ± | ± | ± |
| Langerhans cells | _ | + | + |
| Merkel cells | | | ± |
| Dental Tissue |
| Odontoblasts | | | + |
| Pulp | | | + (mostly due neural elements or monocytes) |
| PDL | | | + (mostly due neural elements or monocytes) |
| Lymphoid tissue |
| Plasma cells | _ | + | |
| Lymphocytes | _ | ± | _ |
| Interdigitating dendritic cells | ± | + | + |
| Follicular dendritic cells | + | + | |
| macrophages | ± | ± | |
| Other connective tissue |
| Striated muscle | ± | _ | ± |
| Schwann cells | _ | + | |
| Vascular endothelium | _ | _ | _ |
| Vascular smooth muscle | _ | ± | |
| Chondrocytes | + | + | |
| Adipocytes | | | ± |
S100 in Oral Squamous Cell Carcinoma (OSCC): S100 proteins in cancer cell differentiation and proliferation: Dysregulation of differentiation is one of the hall marks of human cancer [5]. Most of the S100 proteins are clustered at the chromosomal region 1q21, constituting important components of the epidermal differentiation complex [11]. Therefore, S100 proteins are involved in the terminal differentiation of human epidermis and implicated in cancer and several disorders [5]. Few studies have shown increased S100 protein expression with tumor differentiation whereas others have shown inverse relation. For instance, levels of S100A7 [12], were up regulated but S100A2 [13], S100A6 [14], S100A8 and S100A9 [15,16], S100A14 and S100A16 [17,18] were down regulated with tumor differentiation. Study done by M Roesch-Ely et al., showed S100A 8 and 9 dropped in protein level from tumor distant to tumor adjacent, these signify early changes occurring during field cancerization [19].
Recent study by Sapkota et al., have found S100A14 will modulate the expression of MMP1 and MMP9 in invasive tumor cells of OSCC [20].
S100 and Receptor for Advanced Glycation End product (RAGE) in OSCC: Many evidence have shown, S100 proteins are secreted in extracellular space which act either in autocrine or paracrine mode via common receptor: the RAGE general receptors present in cell surface, implicated in several pathologies including inflammation and cancer [21,22].
In 1992, RAGE was discovered as a cell surface molecule and as a member of immunoglobulin protein because of structural homology with other immunoglobulin protein [22,23]. The mature 382 amino-acid long RAGE is composed of an extracellular part (314 aa), a single transmembrane spanning helix (27aa) and a short cytosolic domain (41aa). The extracellular part of RAGE contains one variable like V-domain (residues 24–127) and two constant like C type domains frequently referred to as C1 (residues 132–230) and C2 domains (residues 239–320). RAGE possesses two N-glycosylation sites, one adjacent to the V-domain and the second one within the V-domain [Table/Fig-3] [21–23].
Structure of RAGE showing relation with various S100 proteins (*indicates that the RAGE domain where S100 binds is not known) [21].
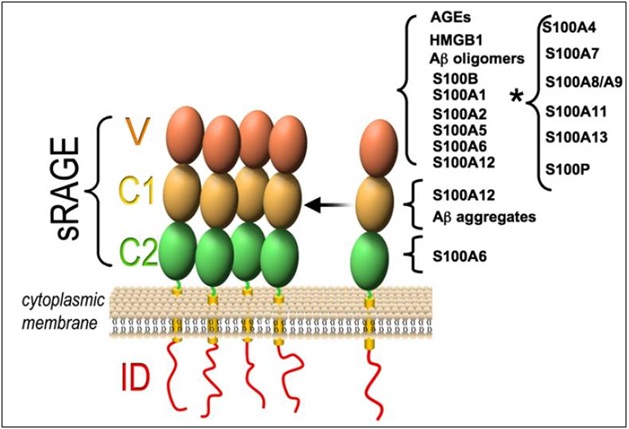
S100A1, S100A2, S100A4, S100A5, S100A6, S100A7, S100A8/A9, S100A11, S100A12, S100A13, S100B, and S100P will interact with RAGE release from cancer cells and [21] activates multiple signaling pathways which includes reactive oxygen species, p21ras, MAP Kinase NF-κB, phosphatidylinositol 3-kinase (PI-3K)/AKT [22]. Thereby S100 is involved in the regulation of diverse cellular processes including inflammation and cancer [5,23].
RAGE is normally present in adult tissue in low levels; but few regions like skin and lung express high levels throughout the life [23]. Expression of RAGE decreases from normal epithelium to OSCC; in normal epithelium it is expressed in spinous cell layer and basal cell layer but few studies have shown that there is frequent absence of expression in basal cell layer [24]. Few studies have shown decreased RAGE expression with tumor differentiation [24, 25]. RAGE was initially identified as receptor for AGE products [26,27]. Besides binding to multiple ligands but also has affinity towards HMGB1 (High Mobility Group Box 1), which is DNA binding protein that functions within the nucleus of cell and also present on surface of many transformed cell lines [23,27].
RAGE-HMGB1 is deeply correlated with tumor progression, metastasis, prognosis of GI cancer, prostate cancer and even plays an important role in OSCC. The interaction between HMGB1 and RAGE stimulates tumor cell proliferation, angiogenesis by activation of VEGF-A, migration and invasion, purportedly through up-regulation of the matrix metalloproteinases MMP-2 and MMP-9, as well as the MAP Kinases [Table/Fig-4] [26,27].
Showing relation of S100 with RAGE and its ligand in tumor progression [28].
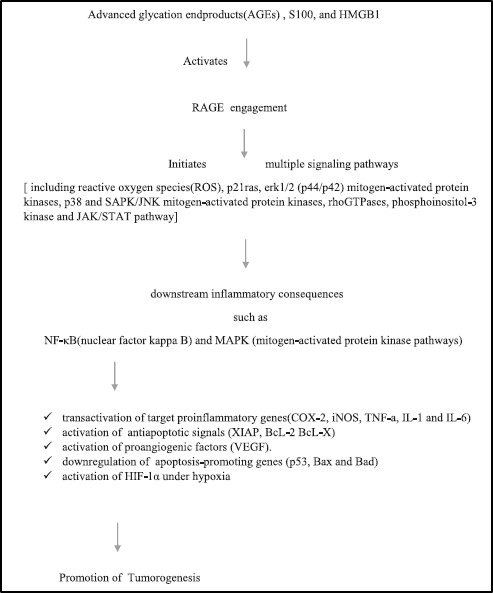
Few studies have shown association of RAGE and OSCC in invasive activity, angiogenesis, prediction of recurrence and nodal metastasis and lymphangiogenesis.
Studies of Landesberg et al., and Sasahira et al., have shown decreased RAGE expression with less differentiated tumor cells, in both NSCLCs and OSCC suggesting their definite etiologic link to tobacco exposure [24]. In the most recent study by Clarissa et al., the first evidence of interaction between HMGB1 and regulatory T cells (Treg), through expression of the HMGB1-recognizing receptors, toll like receptors (TLR4) and RAGE in patients with HNSCC was reported establishing their role in antitumor immune response [28,29].
Recently the use of Etodolac, a selective COX-2 inhibitor has shown significant reduction in the expression of RAGE in dysplasia and OSCC; suggest targeting RAGE-HMGB1 system for OSCC treatment and prognosis [30].
S100 in Granular Cell Tumors: Granular Cell Tumors (GCT) first described by Weber in 1854, are uncommon benign tumours [31]. In 1926, Abrikossoff reported the occurrence of granular cell tumour in the tongue. These tumours are anticipated to originate from striated muscle cells due to histologic similarity to degenerating skeletal muscle and so named as “Myoblastomas” [32,33]. However, soon theory of neural origin was proposed due to close association of tumor with nerve bundles and ganglions. Moreover these tumor cells have shown to react with S100 and other neural markers reinforcing its neural origin [34,35]. Studies have shown strong positivity for S100 in GCT and weak positivity in Congenital Gingival Granular Cell Tumour (CGGCT) but at the same time both the tumors were positive for macrophage markers. Above results propose that, rather than neural origin, granular cells may be derived from pericyte or a related cell with potential smooth muscle differentiation [2]. Study was done by Marilena et al., using panel of IHC markers in oral GCT, showed strong diffuse positivity for p75, vimentin, calretinin and NKI/C3, inhibin-α, PGP9.5, S-100 and CD 68. Thus granular cells possess multifaceted immunoprofile; may also due to stress induced degenerative process or a metabolic disorder they lose their footprint of the cell of origin leading to expression of aberrant and uncharacteristic proteins [36].
S100 in Oral Mucosal Melanoma: Primary Oral Malignant Melanoma (OMM) first reported by Weber in 1856, accounts for only 0.2-0.8% of all malignant melanomas [2] and 0.7% of all mucosal neoplasms in head and neck [37,38]. OMM differs from its cutaneous counterpart in incidence, clinical as well as histologic presentation and it is biologically aggressive with a poor prognosis due to presence of vertical growth phase at the time of diagnosis [2, 39, 40]. OMM shows wide spectrum of histopathological features like epithelioid, spindle, plasmacytoid and clear cell types which mimic epithelial, hematological, mesenchymal and neural tumors, thus making IHC a primary tool to establish the correct diagnosis [41]. Oropharyngeal melanomas have shown inconsistent staining to S100 which has been attributed to lack of S100 DNA in tumor cells or altered S100 profiles as the disease progresses [2]. Few studies showed S100 as a more sensitive marker than HMB-5 and Melan-A in primary OMM [38, 41, 42].
S100 in Salivary Gland Tumors: Salivary gland tumors are heterogeneous group of tumors showing greatest diversity histologically due to presence of myoepithelial cells in various tumors and have major histogenetic implications [43]. Controversy still exists concerning its participation in certain tumors [Table/Fig-5] [44]. Though S100 protein is considered as a traditional marker for myoepithelial cells it is positive only in few studies while negative in other studies in normal salivary gland [45–49]. This incongruity is due to the fact that acini and ducts are associated with rich autonomic nerves which may misconstrue as S100 positive myoepithelial cells or this population of developing myoepithelial cells is heterogeneous and some of them may be representative of a pluripotent population [45,49].
Showing Myoepithelial participation in salivary gland tumors.
| Benign salivary gland tumor | Malignant salivary gland tumor |
|---|
| No MP | Weak MP | More MP | No MP | Weak MP | More MP |
|---|
| Warthin’s tumorOncocytomasDuctal papillomaCanalicular adenoma | Basal cell adenoma | Pleomorphic adenomaMyoepith-elioma | Acinic cell carcinoma,Salivary duct carcinoma,Oncocytic carcinoma | Basal cell adenocar-cinoma,PLGA,MEC | ADCC,Myoepithelial carcinoma,Pleomorphic adenoma ex carcinoma |
MP- Myoepithelial Participation, PLGA- Polymorphus Low Grade Adenocarcinoma, MEC- Mucoepidermoid Carcinoma, ADCC- Adenoid Cystic Carcinoma.
S 100 positivity has been observed in myoepithelial cells of pleomorphic adenoma and myoepithelioma, epithelial cells of canalicular adenoma [46], ductal cells of adenoid cystic carcinoma, epithelial myoepithelial carcinoma, acinic cell adenocarcinoma, polymorphous low grade adenocarcinoma and undifferentiated carcinoma [50,51]. However S100 protein is absent in myoepithelium of normal salivary glands, this reflects acquisition of ectopic proteins in tumorigenesis of salivary glands [2,7].
S100 in Odontogenic Cysts and Tumors: Odontogenic tumors resemble various stages of tooth development, and are derived from epithelial, ectomesenchymal, or both [2]. Ectomesenchymal part of tooth germ is believed to originate from neural crest cells [2,51], thus steered to investigate with S100 protein in one aspect, whereas in other aspect S 100 is investigated due to presence of Langerhan’s Cells (LC) in inflammatory component of connective tissue of odontogenic cyst and tumor [2].
Few studies have showed the presence of LC by S100 positivity in lining epithelia and inflammatory fibrous connective tissue of radicular cyst, dentigerous cyst and odontogenic keratocyst, LC’s distribution seems to be associated with the degree of differentiation of the epithelium [52–54]. Study done by Herald JS on 24 cases of ameloblastomas showed positivity in 10 cases and five cases demonstrated weak positive, this positivity is attributed to neural crest origin, i.e. LC which are usually accompanied with high degree of inflammatory cell infiltrate. The presence of S100 positivity within the ameloblastic strands may be due to presence of LC which supports the hypothesis that ameloblastomas are recapitulation of odontogenic epithelium to the original oral epithelium [55]. S100 positive reaction in granular cells in ameloblastoma is due to presence of lysosomal bodies in those cells [56].
Study by Takeda et al., have shown S100 positivity in both epithelial and ectomesenchymal components of Ameloblastic Fibroma (AF) and ameloblastic fibrodentinoma. This expression of neural protein is related to neural crest related cells in dentinogenesis under certain pathologic conditions in odontogenic tumour [2,53]. Study done by Tadakatsu K et al., showed S100 protein expression in calcified areas in odontogenic tumor suggesting that S100 protein has a biological role in ectopic calcification and may participate in particular Ca2+ signal mechanism in those cells or tissues [56].
S100 in Soft Tissue Tumors: Most of the neural crest derived cells like astrocytes, oligodendrocytes, schwann cells, folliculostellate cells of adenohypophysis, satellite cells of adrenal medulla, chondrocytes, adipocytes, and various histiocytes which include Langerhan’s cells of epidermis and interdigitating reticulum cells of the lymph nodes demonstrate S100 protein and are widely distributed in central and peripheral nervous system [1,7,9,10,51].
Both neurilemoma and neurofibroma show S100 positivity, but intensity and percentage of positive cells are more in neurilemoma due to the uniform population of schwann cells [51,52]. S100 protein staining is vital in the diagnosis of cellular schwannoma, which is often mistaken for fibrosarcoma or low grade Malignant Peripheral Nerve Sheath Tumour (MPNST) [51]. MPNST shows only 50% positivity for S100 which is weak or focal staining. Paragangliomas, neuroblastoma, granular cell tumour show variable expression of S100 [51, 54]. Nerve sheath myxoma shows strong expression of S100 protein [55,56]. Stefansson et al., first identified S100 protein in human adult and fetal chondrocytes, in both cytoplasm and nucleus [7,57].
Subsequently, several groups have identified S 100 protein in normal tissue and in tumors of cartilage like chondroma, chondroblastoma and mesenchymal chondrosarcoma. Earlier, S100 was as a primarily marker for chondrogenic origin. But recently, it has been reported that S100 is involved in cartilage repair process as studies have shown increased expression of S100 in chondrocytes near cartilage lesions [57]. In contrast, benign osseous tumors are S100 negative but cases of osteosarcoma show S100 positivity in the foci of cartilage differentiation [58,59].
Other few soft tissue tumors like Ewing’s Sarcoma / Primitive neuroectodermal tumor, synovial sarcoma, alveolar soft tissue sarcoma, liposarcoma, pleomorphic undifferentiated sarcoma, rhabdomyosarcoma, clear cell sarcoma shows variable S100 positivity [58–60].
Conclusion
S100 protein was first believed to be a neural crest origin but latter showed wider distribution. Due to lack of specificity of S100 protein studies have revealed unclear and often erratic data. Histogenesis of oral and maxillofacial lesions has been investigated with S100 protein, but there interaction with several effector protein are involved in the regulation of diverse spectrum of cellular process. Though few of the S100 proteins have been successfully investigated in clinical context, their implications in pathophysiological process need further clarification.
NSCLC- Non-Small Cell Lung Cancer, OSCC- Oral Squamous Cell Carcinoma, ESCC- Esophagus Squamous Cell Carcinoma, LSCC- Laryngeal Squamous Cell Carcinoma, CRC- Colorectal Cancer.
MP- Myoepithelial Participation, PLGA- Polymorphus Low Grade Adenocarcinoma, MEC- Mucoepidermoid Carcinoma, ADCC- Adenoid Cystic Carcinoma.
[1]. Sedaghat F, Notopoulos A, S100 protein family and its application in clinical practiceHippokratia 2008 12(4):198-204. [Google Scholar]
[2]. Nair SM, Diagnostic and histogenetic significance of s 100 protein in oral diseasesOral Max Path J 2013 4(1):302-08. [Google Scholar]
[3]. Maletzki C, Bodammer P, Breitruck A, Kerkhoff C, S100 proteins as diagnostic and prognostic marker in colorectal and hepatocellular carcinomaHepat Mon 2012 12(10):1-16. [Google Scholar]
[4]. Marenholz I, Lovering RC, Heizmann CW, An update of the S100 nomenclatureBiochimica et Biophysica Acta 2006 1763:1282-83. [Google Scholar]
[5]. Hongyan C, Chengshan Xu, Qing’e J, Zhihua L, S100 protein family in human cancerAm J Cancer Res 2014 4(2):89-115. [Google Scholar]
[6]. Yao R, Davidson DD, Lopez-Beltran A, MacLennan GT, Montironi R, Cheng L, The S100 proteins for screening and prognostic grading of bladder cancerHistol Histopathol 2007 22:1025-32. [Google Scholar]
[7]. Barrett AW, Scully C, S100 protein in oral biology and pathologyJ Oral Mod 1994 23:433-40. [Google Scholar]
[8]. Santamaria-kisie L, Rintala-dempsey AC, Shaw GS, Calcium dependent and independent interactions of the S100 protein familyBiochem. J 2006 396:201-14. [Google Scholar]
[9]. Donato R, Cannon BR, Sorci G, Riuzzi F, Hsu K, Weber DJ, Functions of S100 ProteinsCurr Mol Med 2013 13(1):24-57. [Google Scholar]
[10]. Srikrishna G, Freeze HH. S100 protein family and tumorigenesis. Atlas Genet Cytogenet Oncol Haematol. January 2011. URL: http://AtlasGeneticsOncology.org/Deep/S100ProtFamilyTumorID20092.html [Google Scholar]
[11]. Ingo M, Maja Z, David FF, Claude B, Andreas Z, Dietmar M, Identification of human epidermal differentiation complex (EDC)-encoded genes by subtractive hybridization of entire YACs to a gridded keratinocyte cDNA libraryGenome Research 2000 :341-56. [Google Scholar]
[12]. Kesting MR, Sudhoff H, Hasler RJ, Nieberler M, Pautke C, Klaus-Dietrich W, Psoriasin (S100A7) up-regulation in oral squamous cell carcinoma and its relation to clinicopathologic featuresOral Oncol 2009 45:731-36. [Google Scholar]
[13]. Tsai ST, Jin YT, Tsai WC, Wang ST, Lin YC, Chang MT, S100A2, a potential marker for early recurrence in early-stage oral cancerOral Oncol 2005 41:349-57. [Google Scholar]
[14]. Yang X, Wei KJ, Zhang L, Pan HY, Ye DX, Zhong LP, Decreased expression of S100A6 in oral squamous cell carcinomaOncol Rep 2010 24:479-88. [Google Scholar]
[15]. Regina M. S100-RAGE signaling in the pathogenesis of head and neck squamous cell carcinoma. PhD [dissertation]. Germany. Ruperto-Carola University of Heidelberg; 2013. Available from: Open Access Theses and Dissertations [Google Scholar]
[16]. Tadbir AA, Ashraf MJ, Mehrabani G, S100A9 expression in oral squamous cell carcinomaMiddle-East J. Sci. Res 2013 16(6):775-78. [Google Scholar]
[17]. Sapkota D, Costea DE, Magnus B, Bruland O, Lorens JB, Vasstrand EN, S100A14 inhibits proliferation of oral carcinoma derived cells through G1-arrestOral Oncol 2012 8:219-25. [Google Scholar]
[18]. Sapkota D, Costea DE, Ibrahim SO, Johannessen AC, Bruland O, S100A14 Interacts with S100A16 and regulates its expression in human cancer cellsPLoS ONE 2013 8(9):1-10. [Google Scholar]
[19]. Roesch-Ely M, Nees M, Karsai S, Ruess A, Bogumil R, Warnken U, Proteomic analysis reveals successive aberrations in protein expression from healthy mucosa to invasive head and neck cancerOncogene 2007 26:54-64. [Google Scholar]
[20]. Sapkota D, Bruland O, Costea DE, Haugen H, Vasstrand EN, Ibrahim SO, S100A14 regulates the invasive potential of oral squamous cell carcinoma derived cell-lines in vitro by modulating expression of matrix metalloproteinases, MMP1 and MMP9Eur J Cancer 2011 47:600-10. [Google Scholar]
[21]. Leclerc E, Fritz G, Vetter SW, Heizmann CW, Binding of S100 proteins to RAGE: An updateBiochimica et Biophysica Acta 2009 1793:993-1007. [Google Scholar]
[22]. Rojas A, Figueroa H, Morales E, Fueling inflammation at tumor microenvironment: the role of multiligand/rage axisCarcinogenesis 2010 31(3):334-41. [Google Scholar]
[23]. Rojas A, González I, Morales E, Pérez-Castro R, Romero J, Figueroa H, Diabetes and cancer: looking at the multiligand/ RAGE axisWorld J Diabetes 2011 2(7):108-13. [Google Scholar]
[24]. Landesberg R, Woo V, Huang L, Cozin M, Lu Y, Qu W, The expression of the receptor for glycation endproducts (RAGE) in oral squamous cell carcinomasOral Surg Oral Med Oral Pathol Oral Radiol Endod 2008 105:617-24. [Google Scholar]
[25]. Bhawal UK, Ozaki Y, Nishimura M, Sasahira T, Nomura Y, Sato F, Association of expression of receptor for advanced glycation end products and invasive activity of oral squamous cell carcinomaOncol 2005 69(3):246-55. [Google Scholar]
[26]. Shun-Yao K, Ko HA, Shieh TM, Chang W-C, Chen HI, Chang SS, Cell migration is regulated by AGE-RAGE interaction in human oral cancer cells in vitroPLoS ONE 2015 9(10):1-10. [Google Scholar]
[27]. Takasumi S, Luo Y, Ohmori H, Chihara Y, Fujii K, Sasahira T, Advanced glycation end products (AGE) induce the receptor for AGE in the colonic mucosa of azoxymethane-injected fischer 344 rats fed with a high-linoleic acid and high-glucose dietJ Gastroenterol 2012 47(10):1073-83. [Google Scholar]
[28]. Routray S, RAGE, inflammation and oral cancer: recreating the connexionOral Oncol 2014 50:e58-59. [Google Scholar]
[29]. Clarissa AW, Brandau S, Lotfi R, Mattheis S, Xiang Gu, Lang S, HMGB1 is overexpressed in tumor cells and promotes activity of regulatory T cells in patients with head and neck cancerOral Oncol 2012 48:409-16. [Google Scholar]
[30]. Kirita T, Omura K, Oral Cancer: Diagnosis and therapy. Springer. Tokyo (Japan); 2015. Chapter 3Molecular biology of oral cancer:63-82. [Google Scholar]
[31]. Qureshi NA, Tahir M, Carmichael AR, Granular cell tumour of the soft tissues: a case report and literature reviewInt Semin Surg Oncol 2006 24(3):21-23. [Google Scholar]
[32]. Mona Bi, Khalid A, Afif A, Fatani MI. Granular cell tumor: case reportJ Saudi Society Dermato & Dermatolo Surg 2011 15:25-27. [Google Scholar]
[33]. Sena Costa NC, Bertini F, Carvalho YR, Almedia JD, Rodrigues Cavalcante AS, Granular cell tumor presenting as a tongue nodule: two case reportsJ Med Case Rep 2012 6(56):1-4. [Google Scholar]
[34]. Samson WF, Maomi Li, Expression of calretinin and the alpha-subunit of inhibin in granular cell tumorsAm J Clin Pathol 2003 119:259-64. [Google Scholar]
[35]. Brain H Le, Philip JB, Lewis JE, Kapadia SB, Immunohistochemical assessment of inhibin-a, protein gene product 9.5, S100 protein, CD68, and Ki-67 proliferative index with clinical correlationArch Pathol Lab Med 2004 128:771-75. [Google Scholar]
[36]. Marilena V, William MC, Amos B, Granular cell tumor of the oral cavity: updated immunohistochemical profileJ Oral Pathol Med 2009 38:150-59. [Google Scholar]
[37]. Ashvini P, D’souza J, Oral malignant melanoma: A silent killer?J Indian Soc Periodontal 2011 15(4):425-28. [Google Scholar]
[38]. Suresh B, Subramanyam RV, Aparna V, Sowianya P, Intraoral malignant melanomaNiger Med J 2013 54(4):278-81. [Google Scholar]
[39]. Warszawik-Hendzel O, Morika S, Malgorzata O, Lidia R, Melanoma of the oral cavity: pathogenesis, dermoscopy, clinical features, staging and managementJ Dermatol case Rep 2014 2:60-66. [Google Scholar]
[40]. Ajay K, Ruchi B, Devi SH, Harkanwal PS, Primary oral malignant melanoma: clinicopathological series of four casesDent Res J 2012 9(3):338-44. [Google Scholar]
[41]. De-andrade Bruno AB, Primary oral melanoma: a histopathological and immunohistochemical study of 22 cases of Latin AmericaMed Oral Patol Oral Cir Bucal 2012 17(3):e383-88. [Google Scholar]
[42]. Yu CH, Chen HH, Jeng YM, Wang JT, Wang YP, Liu BY, HMB-45 may be a more sensitive marker than S-100 or melan-A for immunohistochemical diagnosis of primary oral and nasal mucosal melanomasJ Oral Pathol Med 2005 3(9):540-45. [Google Scholar]
[43]. Redder CP, Kandagal VS, Nupura V, Pramod SI, Sharan JS, Sameer A, Myoepithelial cells: current perspectives in salivary gland tumorsClin Cancer Investig J 2013 2:101-05. [Google Scholar]
[44]. Anil RP, Adnan TS, Allen MG, Richard JZ, The myoepithelial immunophenotype in 135 benign and malignant salivary gland tumors other than pleomorphic adenomaArch Pathol Lab Med 1999 123:801-06. [Google Scholar]
[45]. Ianez RF, Buim ME, Coutinho-Camillo CM, Schultz R, Soares FA, Lourenco SV, Human salivary gland morphogenesis: myoepithelial cell maturation assessed by immunohistochemical markersHistopathol 2010 57:410-17. [Google Scholar]
[46]. Bansal AK, Bindal R, Kapoor C, Vaidva S, Singh HP, Current concepts in diagnosis of unusual salivary gland tumorsDent Res J 2012 9:S9-19. [Google Scholar]
[47]. Shaobo Z, Conrad S, Jennifer H, Review and updates of immunohistochemistry in selected salivary gland and head and neck tumorsArch Pathol Lab Med 2015 139:55-66. [Google Scholar]
[48]. Dardick I, Stratis M, Parks WR, DeNardi FG, Kahnt HJ, S-100 protein antibodies do not label normal salivary gland myoepitheliumAm J Pathol 1991 138(3):619-28. [Google Scholar]
[49]. Rasheed FS, Majeed AH, Immunohistochemical expression of actin and S100 in pleomorphic adenoma and mucoepidermoid carcinomaJ Bagh College Dentistry 2011 23(2):51-55. [Google Scholar]
[50]. Nagao T, Sato E, Inoue R, Oshiro H, Takahashi RH, Nagai T, Immunohistochemical analysis of salivary gland tumors: application for surgical pathology practiceActa Histochem Cytochem 2012 45(5):269-82. [Google Scholar]
[51]. Varghese SS, Mathew P, Jose J, Diagnostically relevant molecular markers in head and neck neoplasmsISRN Biomarkers2013:1-6. [Google Scholar]
[52]. Fine SW, McClain SA, Li M, Immunohistochemical staining for calretinin is useful for differentiating schwannomas from neurofibromasAm J Clin Pathol 2004 122:552-59. [Google Scholar]
[53]. Praetorious F, Surgical pathology of the head and neck 2009 3rd edUSAInforma Healthcare:1248Leon Barnes, editor [Google Scholar]
[54]. David SG, Mark G, Malignant peripheral nerve sheath tumorsThe Liddy Shriver Sarcoma Initiative 2006 :1-11. [Google Scholar]
[55]. Safadi RA, Hellstein JW, Diab MM, Hammad HM, Nerve sheath myxoma (Neurothekeoma) of the gingiva: a case report and review of the literatureHead and Neck Pathol 2010 4:242-45. [Google Scholar]
[56]. Bhat A, Narasimha A, Vijaya C, Sundeep VK, Nerve sheath myxoma: report of a rare caseJ Clin Diagn Res 2015 9(4):ED07-09. [Google Scholar]
[57]. Raghunatha RY, S100 proteins in cartilage: role in arthritisBiochim Biophys Acta 2012 1822(4):600-06. [Google Scholar]
[58]. Bahrami A, Truong LD, Ro JY, Undifferentiated tumor: true identity by immunohistochemistryArch Pathol Lab Med 2008 132:326-48. [Google Scholar]
[59]. Heim-Hall J, Yohe SL, Application of immunohistochemistry to soft tissue neoplasmsArch Pathol Lab Med 2008 132:476-89. [Google Scholar]
[60]. Vahini G, A clinicopathological study of soft tissue tumors in correlation with immunohistochemistryJ Dent and Med Sci 2015 14(1):31-40. [Google Scholar]