Introduction
The first trophic (survival and growth promoting) factor was discovered in early 1950’s and named Nerve Growth Factor (NGF). This factor had a substantial effect on sensory and sympathetic neurons [1]. This was followed by the discovery of BDNF in 1982, the second member of “neurotropic” family of neurotropic factors. It was shown that BDNF could actually promote survival of subpopulation of dorsal root ganglion neurons and was subsequently isolated from pig brain [2]. Since then, the list of neurotropic factors have only increased with important one’s being neurotrophin-3 (NT3) [3] and neurotrophin-4/5 (NT-4/5) [4,5] and their function in neuronal survival, neuriteout growth, and synaptic plasticity in the nervous system extensively investigated [6].
BDNF is one of the main factors that supports the survival of neurons [7], maturation [8] and differentiation [9] in the nervous system. The protective properties of BDNF are further accentuated under adverse conditions like cerebral ischemia, hypoglycaemia, glutamatergic stimulation and neurotoxicity [10]. BDNF has been associated with stimulation and growth of neurons from neural stem cells [11]. Furthermore, BDNF protein and mRNA have been investigated and found to be present in a plethora of brain regions including but not limited to, basal forebrain, mesencephalon, hypothalamus, olfactory bulb, cortex, hippocampus, brainstem and spinal cord.
BDNF has been associated with multiple disorders, of which brain disorders remain of utmost importance. Decreased level of BDNF has been linked with multiple neurodegenerative diseases such as Parkinson’s disease [12], Huntington’s disease [13] and Multiple sclerosis [14]. BDNF has also been connected to neuropsychiatric diseases such as degenerative disorders [15] and psychiatric disorders like depression [16], schizophrenia [17] and bipolar disorder [18]. In addition to all that, BDNF has been tied with non-brain related issues such as, energy homeostasis, body weight, obesity, type 2 diabetes mellitus and metabolic syndrome [19].
BDNF Origin, Genetics and Chemical Structure:
BDNF belongs to a family of neurotrophic growth factors along with other members of Nerve Growth Factor (NGF); Neurotrophins-3 (NT-3), NT 4/5 and NT6. The synthesis of BDNF happens in Endoplasmic Reticulum (ER) as a precursor protein called pro BDNF with 32-35 kDa mass, which then moves through the golgi apparatus and Trans-Golgi Network (TGN). These pro isoforms are later cleaved to mature forms followed by dimerization and conversion into receptor ligands. There are different modes of secretion of BDNF, it may be secreted as pro-BDNF, an extracellular protease plasmin may cleave it, or show independent biological effect by interacting with the pan-Neurotrophin Receptor p75NTR and other receptors [20]. At least eight 5’ exons (exon I-VIII) with respective promoters in addition to one 3’ exon (exon IX) encode BDNF in both rodents as well as humans [21]. However, two human specific exons called exon Vh and VIIIh have been identified with exon Vh having a specific promoter linked to it, whereas exon VIIIh has no such independent promoter. Therefore, a total of ten exons coding for 5’ untranslated region, which are alternatively spliced to a common 3’ coding exon, have been found in human BDNF gene [21]. BDNF shares about 50% amino acid similarity and structural homology with NGF, NT-3, NT-4/5. Each neurotrophin consists of an initiation codon and pro-region containing an N-linked glycosylation site that is followed by a non-covalently linked homodimer with a signal peptide [22]. Prohormone convertases, such as furin cleave the proneurotrophins (M.W. ~30 kDa) that are initially produced to mature neurotrophin (M.W. ~14kDa) [23]. ([Table/Fig-1]. Structure of BDNF; PDBID-1B8M).
Physiological Effects of BDNF
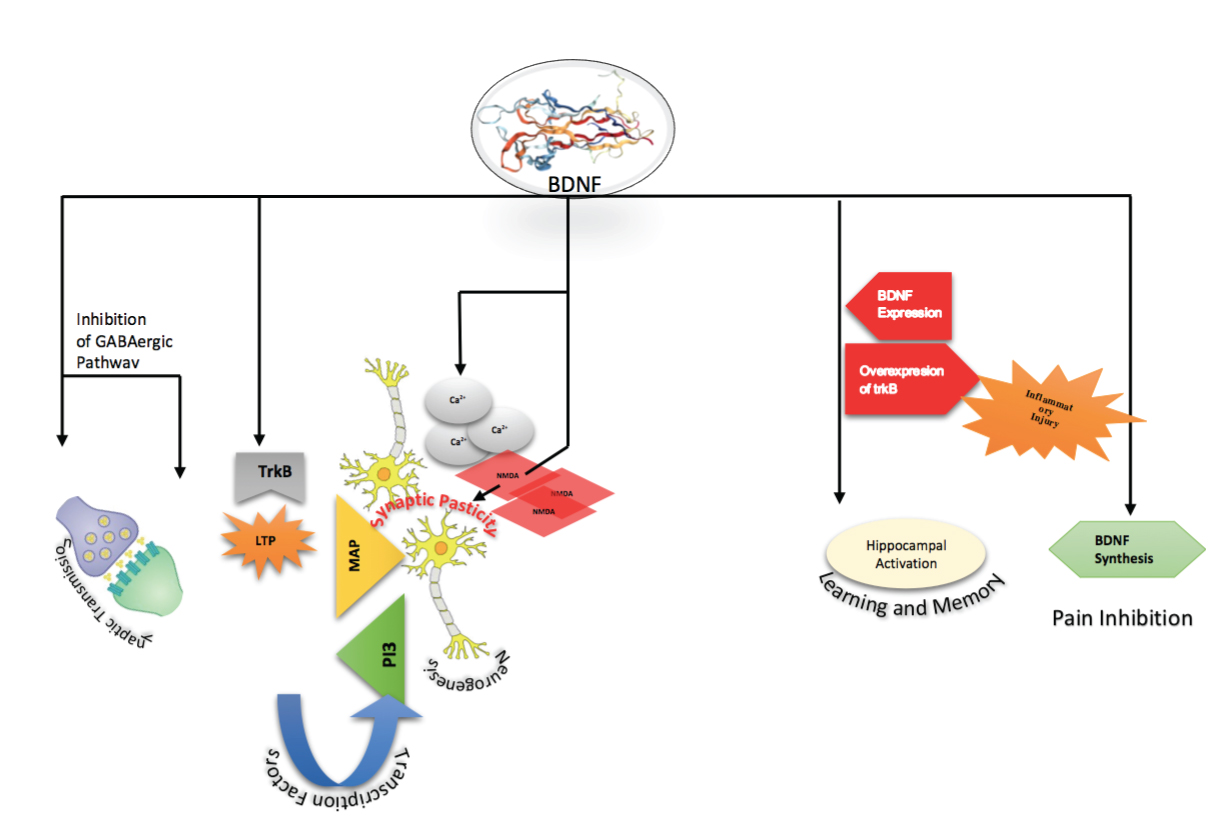
Physiological Role of BDNF
Synaptic Transmission:
During the very first studies on BDNF, it was found that BDNF increased the frequency of miniature Excitatory Postsynaptic Currents (EPSCs) in Xenopus cultures. These studies were eventually followed by numerous independent studies confirming other actions of BDNF in synaptic transmission. These actions include, strengthening of excitatory (glutamatergic) synapses and weakening of inhibitory (GABAergic) synapses. Effect of BDNF on Long-Term Potentiation (LTP) has been supported by various studies among which Schuman et al., prominently demonstrated the exposure of adult rat hippocampal slices to BDNF that leads to long term potentiation of afferent input to hippocampal pyramidal cells [24]. Also, similar studies found that, hippocampal slices from BDNF knockout animals exhibited impaired LTP, which is restored by BDNF reintroduction, further strengthened the original findings [25].
There is, however still huge debate on whether this LTP due to BDNF is primarily a result of presynaptic action, or postsynaptic modifications [26]. This is because a number of studies have supported both, the presynaptic locus [27] as well as postsynaptic actions. Pertinently, both pre- and postsynaptic trkB located in hippocampus might have substantial contribution [28].
Preliminary study probes into influence of BDNF on GABAergic neuronal phenotype raised an interest in role of BDNF in GABAergic synapses. This was soon followed by studies that showed decrease in inhibitory (GABAergic) synaptic transmission in response to BDNF [29]. This action can be partly associated with modulation of GABA receptor phosphorylation [30]. Recently, differential effects of BDNF on GABA-mediated currents in inhibitory and excitatory neuron subpopulations were elucidated, and a selective decrease in efficacy of inhibitory neurotransmission by downregulation of Cl-transport was reported [29].
Neurogenesis:
An important role of BDNF has also been established in neurogenesis. This has been done through various techniques, crucially the adenoviral induced activity and intraventricular infusion of BDNF that accentuates the number of neurons in the adult striatum, septum, thalamus and olfactory bulb [31], which can be potentiated by concurrent inhibition of glial differentiation of subependymal progenitor cells [32]. Further mechanistic investigations on progenitor cells point out to different signaling cascade which appear to involve trkB activation followed by activation of MAP kinase and PI3-kinase pathways, and downstream modification of basic helix-loop-helix transcription factors [33]. In addition to the role of BDNF in neurogenesis primarily through proliferation, other experiments validate BDNF’s effect on survival of neurons with effects also depending on previous history of ischaemic disease [34].
Synaptic Plasticity:
Pre- and post-synaptic mechanisms have both been found to be involved in regulation of activity-dependent synaptic plasticity [31]. In cultured neocortical neurons of BDNF-knockout mice, BDNF is essential for pre-synaptic vesicle cycling and in turn dependent on NMDA (N-methyl D-aspartate) receptor activation [33]. This initial research was further strengthened by confirming the paracrine (retrograde messenger) role of BDNF, and it was shown that the application of BDNF to hippocampal sections restored LTP stability and spine actin polymerisation in rats [35]. In addition, increase in BDNF levels not only prompted the increase in intracellular calcium concentration and NMDA levels, but also relieved Mg2+ block of NMDA receptors [36], eventually leading to promotion of long-term changes in synaptic activity. LTP induction was also found to be downregulated with a decrease in TrkB and BDNF secretion. Therefore, the overall effect of BDNF on synaptic plasticity is mediated through NMDA trafficking by an increase in calcium influx, leading to post-synaptic BDNF release that [37], in-turn increases pre-synaptic vesicle cycling, thereby enhancing LTP and eventually synaptic plasticity [38].
Learning and Memory:
BDNF has been found to be responsible for activity dependent synaptic plasticity, an increased interest has arisen into studying its role in learning and memory. The main site of action of BDNF has been reported to be hippocampus, the part of brain that is mostly associated with long-term memory in humans and animals. This effect has been proven by a number of studies, including the demonstration of rapid and selective induction of BDNF expression in the hippocampus during contextual learning [39], and function-clocking antibodies to BDNF, BDNF knockout, knockout of forebrain trkB signaling, and an impair in spatial learning in mice due to overexpression of truncated trkB [40]. A tool-use learning experiment in monkey revealed an upregulation of BDNF in parietal cortex. In humans, a valine to methioninepolymorphism at the 5′ proregion of the human BDNF protein was found to be associated with poorer episodicmemory in vitro, neurons transfected with met BDNFGFP exhibited reduced depolarisation induced BDNF secretion [41].
Pain:
BDNF plays a major role in pain transduction through its neuromodulatory properties. A marked upregulation and synthesis of BDNF occurs in dorsal horn neurons and in response to inflammatory injury. BDNF inhibitors abrogate the sensitisation of nociceptive afferents and hyperplasia caused by acute BDNF secretion [42]. Behavioural and electrophysiological data demonstrate that BDNF signal transduction inhibits central pain sensitisation, a condition that is caused by activity-dependent increase in excitability of dorsal horn neurons leading to clinical condition named “neuropathic pain” and symptomised by lowering of pain threshold [43].
Endothelial and Cardiac Cells:
BDNF uses signaling through TrkB receptors to promote neovascularisation therapeutically, which is different from the apoptosis induction and angiogenesis repudiation that otherwise occurs through the low-affinity p75 NTR receptor. Involvement of BNDF and NT3 in the formation of myocardial and heart vasculature has been reported through studies carried out in murine BDNF knockout models. This has been linked to two major pathways of TrkB receptors, that is, PI3Kinase/AKT and ERK/MAPK that eventually promote EC survival [44]. Furthermore, the cardiovascular protective properties of BDNF may be attributed to the activation of endothelial Nitric Oxide (NO) synthase through AKT initiation [45].
Lipid Metabolism:
In early 1990’s, it was reported that intracerebroventral (ICV) administration of BDNF in rats affected energy metabolism, eventually leading to decreased energy intake and body weight loss. These actions of BDNF were correlated with dose-dependent increase in serotonin turnover, adaptive plasticity and nerve cell survival [46]. A positive correlation of BDNF with lipid profile has also been established. Studies on diabetic animals that were treated with BDNF also showed promising results with decrease of plasma glucose, non-esterified fat, phospholipids, and liver weight along with an increase in β-oxidation, peroxisome Proliferator Activator Receptor (PPARα) activation and level of fibroblast growth factor [47].
Immunity and Inflammation:
Bronchial Hyperactivity (BHR), a hallmark of allergic asthma has been hypothesised to be due to elevated levels of Neurotrophins (NTs), confirmed from the observation that release of NTs occurred from immune cells including B-lymphocytes, eosinophils, mast cells and macrophages [47]. A hypothesis has been put forward, where the role of BDNF in allergic asthma has been associated with mucus hyper-secretion and enhanced airway smooth muscle contraction resulting due to facilitation of acetyl choline release and extravasation of the plasma. In contrast, because of immunomodulatory action of BDNF, its increased production in neuroinflammatory diseases like multiple sclerosis suggests its neuroprotective activity. All this evidence points out BDNF as a promising therapeutic target for detection and prevention of neurological inflammatory disorders [48].
Clinical Uses of BDNF in Various Disorders:
Bipolar Disorder:
Bipolar Disorder (BD) is a highly chronic mood disorder characterised by the presence of manic and depressive symptoms and a lifetime prevalence of 3.9%. Recent neuropathological studies suggest that BD is caused due to changes in neuronal plasticity, particularly in cell resilience and connectivity. And BDNF has been reported as a major contributor to the neuroplasticity changes described among BD patients. BDNF serum levels have also been reported to decrease in patients with BD that return to normal levels in euthymia. Other non-pathological factors, like life stress and trauma have also been associated with a decreased BDNF serum levels [49]. All these studies suggest BDNF as an important factor in BD and moreover a promising biomarker for diagnosis of bipolar disorder.
Epilepsy:
After the discovery of the role of increased NGF mRNA levels in limbic seizures, it was suggested that seizure-induced expression of neurotrophic factors may contribute to the lasting structural and functional underlying epileptogenesis changes [50]. The electrophysiological and behavioural changes conducted both in vitro and in vivo implicate BDNF as a mediator in the epileptic cascade. Upregulation of BDNF mRNA and protein in brain hippocampus by seizure activity in animal models, infusion of anti-BDNF agents or use of BDNF knockout or the elipleptogenesis inhibition in animal models with truncated trkB-overexpression [51]. Paradoxically, direct application of BDNF induces hyperexcitability in vitro, while in transgenic mice spontaneous seizures are invoked by overexpression of BDNF, and sufficient seizure induction in vivo has been achieved by infusion of BDNF through intrahippocampal route. This pro epileptogenic effect of BDNF has been closely associated with hippocampus and limbic structures, with supporting evidence of increased BDNF expression in the hippocampus specimens from patients with temporal lobe epilepsy [52]. All this evidence points out to BDNF as an effective target for novel anticonvulsant and antiepileptogenic therapies [53].
Alzeihmer’s Disease:
Alzeihmer’s Disease (AD) is the most common age related neurodegenerative disorder with noticeable impairment of cognitive function. A progressive loss of neurons and synapses, which causes loss of new memory formation, has been known, especially in the entorhinal cortex and hippocampus [54]. A decreased level of BDNF protein and mRNA in serum [55] and many parts of brain, importantly hippocampus and cortex of AD patients has been reported [56]. Recently, a mouse model study on animal models of AD confirmed that BDNF gene delivery to entorhinal cortex alleviated entorhinal cortical and hippocampal neuronal degeneration [57]. Other mouse models of AD with amyloid-β overproduction caused by Amyloid Precursor Protein (APP) mutations exhibited significant reduction in cortical BDNF Mrna [58]. Furthermore, BDNF secreted by Neuronal Stem Cell (NSC) after transplantation also improved cognitive function in AD mice model [59], therefore, suggesting BDNF based therapy a promising lead for AD.
Huntington’s Disease:
Huntington’s Disease (HD) is a disease caused by abnormal htt proteins with polyglutamate expansion (polQ), resulting due to repeats of CAG trinucleotide in the Huntington (htt) gene. It is a neurodegenerative and autosomal dominant disease and degeneration of striatal neurons is believed to induce cognitive decline and progressive psychiatric and motor decline [60]. A possible link between HD and BDNF has been reported on the basis of the fact that substantial increase in BDNF levels are found in adult rodent straitum while TrkB mRNA without the presence of BDNF mRNA. This observation is further demonstrated by the fact that cortical neurons projecting to the straitum contain high levels of BDNF mRNA, and most striatal BDNF in anterogradely transported from the cortex. While the non-mutant form (polyQ) could not stimulate transcription, a positive correlation was found with the wild-type htt that stimulated transcription from BDNF exon II in the cereberal cortex [61]. Post-mortem of patient brains with HD revealed a decreased level of BDNF mRNA and protein levels in caudate-putamen, cortex, striatum, cerebellum, and substantia nigra [62]. In addition, a decreased level of serum BDNF was also reported [63]. In mouse models of HD impaired motor function, cognitive decline and premature death was observed [64]. Findings also suggest that mutations in htt lead to decreased amounts of BDNF in the striatum by inhibiting both BDNF transcription and axonal transport of BDNF containing-vesicles in cortical neurons. Furthermore, severely reduced BDNF levels in R6/1 HD mice hippocampus and striatum were rescued by environmental enrichment [64]. The enrichment, consisting of small cardboard boxes, small open wooden boxes, cylindrical cardboard tunnels, and folded sheets of paper prevented body weight loss and ameliorated motorsymptoms of the mice [65]. These studies suggest that interaction between htt and BDNF are a major source of interaction and therefore, TrkB agonists and exogenous supplementation of BDNF would be a primary choice for HD treatment in future.
Major Depressive Disorder:
Major Depressive Disorder (MDD) is a leading cause of disability worldwide and is clinically presented with symptoms including anxiety, inappropriate guilt, anhedonia, appetite change and sleep disturbance [66]. Stressful events like early life trauma as well as acute stress correlate strongly with occurrence of MDD and interestingly enough, BDNF is reduced by stress, an important risk factor in MDD. The correlation between BDNF and MDD is further strengthened by findings that show BDNF levels are increased by anti-depressant treatment. The increasing evidence in support of BDNF as a conduit in MDD has lead investigators to focus on BDNF as a biomarker and also as a potential target for treatment of MDD [64].
Schizophrenia:
Schizophrenia affects 1% of population and is characterised by three major psychiatric manifestations: negative symptoms (lack of pleasure and ability to begin and sustain planned activities (flat affect), positive symptoms (delusions, thought and movement disorders and haellucinations), and cognitive symptoms (impaired ability to understand and use information, and problems with working memory) [67]. It is however observed that the overall number of neurons in prefrontal cortex of patients with schizophrenia is not decreased. Instead, reduced dendritic spine density of pyramidal cells and synaptophysin (presynaptic protein) was observed in cortex, hinting at role of synaptic dysfunction in the pathogenesis of the disease. Expression levels of BDNF or TrkB have been investigated in the brains of schizophrenic patients but whether or not BDNF levels show discrepancies in serum or brain tissue is still controversial [68]. There is still debate on presence of BDNF in certain brain regions, with certain studies showing increased levels of BDNF in hippocampal and cortical tissues while other studies show a decreased level of BDNF in these brain regions [69]. Furthermore, reduced level of BDNF protein and mRNA have been consistently reported over the course of time, with some reports showing an increased serum BDNF mRNA levels in schizophrenic patients treated with Clozapine. Phencyclidine (PCP), MK-801, or ibotenic acid induced schizophrenia in animal models have shown decreased BDNF mRNA and protein levels. On the other hand, second generation anti psychotics like risperidone and olanzapine in-addition to first generation antipsychotics such as haloperidol and chlorpramazine, all tend to reduce BDNF protein levels in the rat hippocampus, straitum and cortex [70]. A consistency of observation has been reported with demonstration of reduced cortical synaptic structure in cortical neurons and as a result of decreased BDNF secretion [71]. Based on current resaerch, there is therefore, insufficient information to deduce whether a BDNF based approach would be fitting as schizophrenia treatment.
Anxiety Related Disorders:
Anxiety-related disorders in humans include generalised anxiety disorder as well as phobias, panic disorder, obsessive compulsive disorders, and Post-Traumatic Stress Disorders (PTSD) [72]. Role of BDNF in anxiety has been thoroughly researched and a few conclusions have been drawn from the research. In conditional mutants for BDNF mutant female micelines, where the gene was deleted in the broad forebrain, decreased anxiety was reported [73]. A difference in gender anxiety was also note when female mice showed greater susceptibility after stress for enhanced anxiety than stressed mutant male mice [74]. Also, homozygous Mice for V66M allele show an increase in anxiety related behaviour [75]. However, in humans mixed results were found in relation to V66M allele. While some studies show an enhanced effect of V66M SNP on anxiety-levels, meta-analysis shows no such substantial effect [76]. However, a recent study has demonstrated impairments in fair extinction response in human V66M carriers and therefore a link between BDNF and anxiety-related disorders can be modestly framed [77].
Post Traumatic Stress Disorder (PTSD):
Studies have been conducted to find out the role of BDNF in abnormal fear memory and extinction [78] and since pathological fear in a central feature of PTSD, BDNF gene serves as an interesting strarting point for research into its role in PTSD. With that basic idea in mind, rodent studies were carried out where V66M mutations were correlated with impaired fear memory [75], similar to findings with another BDNF mouse model [79]. A loss of fear memory acquisition was induced in targeted knockdown or inhibition of BDNF TrkB signalling in the amygdala [80]. Morevover, hippocampal deletetion of BDNF leads to disrupted extinction of fear memory [81]. Taken together, these studies confirm a possible role of BDNF in PTSD and thus, a possible therapeutic target for PTSD and especially for fear memory formation and extinction.
Type II Diabetes Mellitus:
Role of BDNF in the regulation of food intake and body weight has been scienitifically established both in experimental animals as well as humans. In one study, it was found that systemic administration of BDNF decreased non-fasted blood glucose in obese, non-insulin dependent diabetic mice with an collateral decrease in the body weight [82]. BDNF was found to help in blood glucose control, which was however not associated with decreased food intake but instead to reduced liver glycogen and liver enzyme activity in serum, supporting the liver based mechanism of action of BDNF in blood glucose control [83].
Further support for the role of BDNF in diabetes mellitus is evident from the observation that once or twice perweek administration (70 mg/kg/wk) to db/db mice for three weeks significantly reduced blood glucose concentration and haemoglobin A (HbA) compared to controls. These results suggest that BDNF not only reduced blood glucose concentrations but also restored systemic glucose balance, hinting at the possibility that BDNF could be a novel hypoglycaemic agent even with treatment as infrequently as once per week [84]. Moreover, a lowered blood glucose level, increased uncoupling protein-1 mRNA expression, increased pancreatic insulin content and enhanced thermogenesis and norepinephrine turnover was reported in db/db mice upon Intracerbroventricular (ICV) administration of BDNF [85].
Human studies further strengthens the role of BDNF in type II diabetes where a decrease in plasma levels of BDNF were reported, independent of obesity, and inversely associated with fasting plasma blood glucose, but not with insulin [86]. When output of BDNF from the human brain was studied, output was inhibited when blood glucose levels were elevated, whereas when plasma insulin was increased while maintaining normal blood glucose, the cerebral output of BDNF was not inhibited. These results indicate that high levels of glucose, but not insulin, inhibited the output of BDNF from the human brain. These results emphasise that low levels of BDNF accompany impaired glucose metabolism, and decreased BDNF may be a factor involved in type 2 diabetes mellitus [87].
Blindness and Deafness:
Blindness and deafness are partly neurodegenerative disseases that effects millions of people worldwide [88]. In blindness, almost all the factors finally lead to Retinal Ganglionic Cells (RGCs) neurodegeneration and subsequent loss. A promising positive effect of BDNF on survival of RGC has been established in vitro [89]. Moreover, at the retina, BDNF receptor, TrkB, has been found to be expressed by a number of cell types, namely, a subset of cone photoreceptors, RGCs, Muller glia, and amacrine cells [90]. But the problem with blindness and deafness is that delivery of BDNF becomes a lot more daunting considering the blood-retinal and blood-cochlear barrier, but a lot of efforts have been made for better delivery of BDNF to the targeted site [91], therefore, making it one of the best therapeutic option for both blindness as well as deafness.
Factors that Increase BDNF Levels:
Considering the positive role of increased levels of BDNF in a plethora of diseases and disorders listed in this review, a lot of interest has grown into determining and studying factors that increase BDNF levels in brain and other organs. Chemical substances that increase BDNF include, 7,8-dihroxyflavone, ampakines, LM22A-4, thyroid hormone, melatonin, intranasal insulin, intranasal oxytocin, testosterone, progesterone, adenosine, nicotine, resveratrol, folic acid, piperine, and zinc. Natural BDNF enhancers include Panax ginseng, Omega-3, quercitin, nobiletin, Vitamin D, blueberries, cocao or dark chocolate, and curcumin. Behavioural and physical factors that increase the levels of BDNF include, enriched housing, socialisation, meditation, calorie restriction, sex, sunlight, exercise, bright-light therapy, deep-brain stimulation, acupuncture, electroconvulsive therapy and high-frequency repetitive Transcranial Magnetic Stimulation (rTMS) [92].
Epigenetics of BDNF:
Promoter methylation has been suggested by many studies to be responsible for the regulation of BDNF expression. Equally surmountable experiments point out to the possibility that histone modifications at the promoter region of BDNF might underlie a myriad of neurological pathological processes [93]. Therefore, a robust study on various epigenetic BDNF factors might help scientists to better understand pathology and molecular mechanisms of various diseases and disorders and eventually lay a fitting foundation for future therapeutics.
Conclusion
A thorough literature review on BDNF revealed an immense impact of BDNF in both normal physiology as well as pathology of various diseases ranging right from neurodegenerative and neuropsychiatric to metabolic and sensory disorders. A lot of research suggests BDNF, its increase in some parts of brain regions and other organs as a possible future therapeutic target and this review should serve as a starting point and an important reference for most of the promising BDNF targets in different clinical diseases and disorders in future.
[1]. Levi-Montalcini R, Hamburger V, Selective growth stimulating effects of mouse sarcoma on the sensory and sympathetic nervous system of the chick embryoJournal of Experimental Zoology Part A: Ecological Genetics and Physiology 1951 116(2):321-61.10.1002/jez.1401160206 [Google Scholar] [CrossRef]
[2]. Barde Y-A, Edgar D, Thoenen H, Purification of a new neurotrophic factor from mammalian brainThe EMBOJournal 1982 1(5):54910.1002/j.1460-2075.1982.tb01207.x7188352 [Google Scholar] [CrossRef] [PubMed]
[3]. Maisonpierre PC, Belluscio L, Squinto S, Ip NY, Furth ME, Lindsay RM, Neurotrophin-3: a neurotrophic factor related to NGF and BDNFScience 1990 247(4949):1446-52.10.1126/science.23210062321006 [Google Scholar] [CrossRef] [PubMed]
[4]. Hallböök F, Ibáñez CF, Persson H, Evolutionary studies of the nerve growth factor family reveal a novel member abundantly expressed in Xenopus ovaryNeuron 1991 6(5):845-58.10.1016/0896-6273(91)90180-8 [Google Scholar] [CrossRef]
[5]. Ip NY, Ibanez CF, Nye SH, McClain J, Jones PF, Gies DR, Mammalian neurotrophin-4: structure, chromosomal localization, tissue distribution, and receptor specificityProceedings of the National Academy of Sciences 1992 89(7):3060-64.10.1073/pnas.89.7.30601313578 [Google Scholar] [CrossRef] [PubMed]
[6]. van Velthoven CT, Sheldon RA, Kavelaars A, Derugin N, Vexler ZS, Willemen HL, Mesenchymal stem cell transplantation attenuates brain injury after neonatal strokeStroke 2013 44(5):1426-32.10.1161/STROKEAHA.111.00032623539530 [Google Scholar] [CrossRef] [PubMed]
[7]. Huang EJ, Reichardt LF, Neurotrophins: roles in neuronal development and functionAnnual Review of Neuroscience 2001 24(1):677-736.10.1146/annurev.neuro.24.1.67711520916 [Google Scholar] [CrossRef] [PubMed]
[8]. Acheson A, Conover JC, Fandl JP, DeChiara TM, Russell M, Thadani A, A BDNF autocrine loop in adult sensory neurons prevents cell deathNature 1995 374(6521):450-53.10.1038/374450a07700353 [Google Scholar] [CrossRef] [PubMed]
[9]. Vivar C, Potter MC, van Praag H, All about running: synaptic plasticity, growth factors and adult hippocampal neurogenesisNeurogenesis and Neural Plasticity 2012 Springer:189-210.10.1007/7854_2012_22022847651 [Google Scholar] [CrossRef] [PubMed]
[10]. Maisonpierre PC, Le Beau MM, Espinosa R, Ip NY, Belluscio L, Suzanne M, Human and rat brain-derived neurotrophic factor and neurotrophin-3: gene structures, distributions, and chromosomal localizationsGenomics 1991 10(3):558-68.10.1016/0888-7543(91)90436-I [Google Scholar] [CrossRef]
[11]. Benraiss A, Chmielnicki E, Lerner K, Roh D, Goldman SA, Adenoviral brain-derived neurotrophic factor induces both neostriatal and olfactory neuronal recruitment from endogenous progenitor cells in the adult forebrainJournal of Neuroscience 2001 21(17):6718-31.10.1523/JNEUROSCI.21-17-06718.200111517261 [Google Scholar] [CrossRef] [PubMed]
[12]. Scalzo P, Kümmer A, Bretas TL, Cardoso F, Teixeira AL, Serum levels of brain-derived neurotrophic factor correlate with motor impairment in Parkinson’s diseaseJournal of Neurology 2010 257(4):540-45.10.1007/s00415-009-5357-219847468 [Google Scholar] [CrossRef] [PubMed]
[13]. Mughal MR, Baharani A, Chigurupati S, Son TG, Chen E, Yang P, Electroconvulsive shock ameliorates disease processes and extends survival in huntingtin mutant miceHuman Molecular Genetics 2010 20(4):659-69.10.1093/hmg/ddq51221106706 [Google Scholar] [CrossRef] [PubMed]
[14]. Sohrabji F, Lewis DK, Estrogen–BDNF interactions: implications for neurodegenerative diseasesFrontiers in neuroendocrinology 2006 27(4):404-14.10.1016/j.yfrne.2006.09.00317069877 [Google Scholar] [CrossRef] [PubMed]
[15]. Altar CA, Neurotrophins and depressionTrends in Pharmacological Sciences 1999 20(2):59-62.10.1016/S0165-6147(99)01309-7 [Google Scholar] [CrossRef]
[16]. Duman RS, Monteggia LM, A neurotrophic model for stress-related mood disordersBiological Psychiatry 2006 59(12):1116-27.10.1016/j.biopsych.2006.02.01316631126 [Google Scholar] [CrossRef] [PubMed]
[17]. Favalli G, Li J, Belmonte-de-Abreu P, Wong AH, Daskalakis ZJ, The role of BDNF in the pathophysiology and treatment of schizophreniaJournal of Psychiatric Research 2012 46(1):1-11.10.1016/j.jpsychires.2011.09.02222030467 [Google Scholar] [CrossRef] [PubMed]
[18]. Barnett JH, Smoller JW, The genetics of bipolar disorderNeuroscience 2009 164(1):331-43.10.1016/j.neuroscience.2009.03.08019358880 [Google Scholar] [CrossRef] [PubMed]
[19]. Bathina S, Das UN, Brain-derived neurotrophic factor and its clinical implicationsArchives of medical science: AMS 2015 11(6):116410.5114/aoms.2015.5634226788077 [Google Scholar] [CrossRef] [PubMed]
[20]. Teng HK, Teng KK, Lee R, Wright S, Tevar S, Almeida RD, ProBDNF induces neuronal apoptosis via activation of a receptor complex of p75NTR and sortilinJournal of Neuroscience 2005 25(22):5455-63.10.1523/JNEUROSCI.5123-04.200515930396 [Google Scholar] [CrossRef] [PubMed]
[21]. Pruunsild P, Kazantseva A, Aid T, Palm K, Timmusk T, Dissecting the human BDNF locus: bidirectional transcription, complex splicing, and multiple promotersGenomics 2007 90(3):397-406.10.1016/j.ygeno.2007.05.00417629449 [Google Scholar] [CrossRef] [PubMed]
[22]. Klein R, Nanduri V, Jing S, Lamballe F, Tapley P, Bryant S, The trkB tyrosine protein kinase is a receptor for brain-derived neurotrophic factor and neurotrophin-3Cell 1991 66(2):395-403.10.1016/0092-8674(91)90628-C [Google Scholar] [CrossRef]
[23]. Chao MV, Bothwell M, Neurotrophins: to cleave or not to cleaveNeuron 2002 33(1):9-12.10.1016/S0896-6273(01)00573-6 [Google Scholar] [CrossRef]
[24]. Kang H, Schuman EM, Long-lasting neurotrophin-induced enhancement of synaptic transmission in the adult hippocampusScience 1995 267(5204):1658-62.10.1126/science.78864577886457 [Google Scholar] [CrossRef] [PubMed]
[25]. Korte M, Griesbeck O, Gravel C, Carroll P, Staiger V, Thoenen H, Virus-mediated gene transfer into hippocampal CA1 region restores long-term potentiation in brain-derived neurotrophic factor mutant miceProceedings of the National Academy of Sciences 1996 93(22):12547-52.10.1073/pnas.93.22.125478901619 [Google Scholar] [CrossRef] [PubMed]
[26]. Schinder AF, Poo M-m, The neurotrophin hypothesis for synaptic plasticityTrends in Neurosciences 2000 23(12):639-45.10.1016/S0166-2236(00)01672-6 [Google Scholar] [CrossRef]
[27]. Kafitz KW, Rose CR, Thoenen H, Konnerth A, Neurotrophin-evoked rapid excitation through TrkB receptorsNature 1999 401(6756):918-21.10.1038/4484710553907 [Google Scholar] [CrossRef] [PubMed]
[28]. Drake C, Milner T, Patterson S, Ultrastructural localization of full-length trkB immunoreactivity in rat hippocampus suggests multiple roles in modulating activity-dependent synaptic plasticityJournal of Neuroscience 1999 19(18):8009-26.10.1523/JNEUROSCI.19-18-08009.199910479701 [Google Scholar] [CrossRef] [PubMed]
[29]. Wardle RA, Poo M-m, Brain-derived neurotrophic factor modulation of GABAergic synapses by postsynaptic regulation of chloride transportJournal of Neuroscience 2003 23(25):8722-32.10.1523/JNEUROSCI.23-25-08722.200314507972 [Google Scholar] [CrossRef] [PubMed]
[30]. Jovanovic JN, Thomas P, Kittler JT, Smart TG, Moss SJ, Brain-derived neurotrophic factor modulates fast synaptic inhibition by regulating GABAA receptor phosphorylation, activity, and cell-surface stabilityJournal of Neuroscience 2004 24(2):522-30.10.1523/JNEUROSCI.3606-03.200414724252 [Google Scholar] [CrossRef] [PubMed]
[31]. Pencea V, Bingaman KD, Wiegand SJ, Luskin MB, Infusion of brain-derived neurotrophic factor into the lateral ventricle of the adult rat leads to new neurons in the parenchyma of the striatum, septum, thalamus, and hypothalamusJournal of Neuroscience 2001 21(17):6706-17.10.1523/JNEUROSCI.21-17-06706.200111517260 [Google Scholar] [CrossRef] [PubMed]
[32]. Chmielnicki E, Benraiss A, Economides AN, Goldman SA, Adenovirally expressed noggin and brain-derived neurotrophic factor cooperate to induce new medium spiny neurons from resident progenitor cells in the adult striatal ventricular zoneJournal of Neuroscience 2004 24(9):2133-42.10.1523/JNEUROSCI.1554-03.200414999064 [Google Scholar] [CrossRef] [PubMed]
[33]. Ito H, Nakajima A, Nomoto H, Furukawa S, Neurotrophins facilitate neuronal differentiation of cultured neural stem cells via induction of mRNA expression of basic helix-loop-helix transcription factors Mash1 and Math1Journal of Neuroscience Research 2003 71(5):648-58.10.1002/jnr.1053212584723 [Google Scholar] [CrossRef] [PubMed]
[34]. Gustafsson E, Lindvall O, Kokaia Z, Intraventricular infusion of TrkB-Fc fusion protein promotes ischemia-induced neurogenesis in adult rat dentate gyrusStroke 2003 34(11):2710-15.10.1161/01.STR.0000096025.35225.3614563966 [Google Scholar] [CrossRef] [PubMed]
[35]. Walz C, Jüngling K, Lessmann V, Gottmann K, Presynaptic plasticity in an immature neocortical network requires NMDA receptor activation and BDNF releaseJournal of Neurophysiology 2006 96(6):3512-16.10.1152/jn.00018.200617110740 [Google Scholar] [CrossRef] [PubMed]
[36]. Kramár EA, Chen LY, Lauterborn JC, Simmons DA, Gall CM, Lynch G, BDNF upregulation rescues synaptic plasticity in middle-aged ovariectomized ratsNeurobiology of Aging 2012 33(4):708-19.10.1016/j.neurobiolaging.2010.06.00820674095 [Google Scholar] [CrossRef] [PubMed]
[37]. Ellis-Davies GC, Two-photon microscopy for chemical neuroscienceACS Chemical Neuroscience 2011 2(4):18510.1021/cn100111a21731799 [Google Scholar] [CrossRef] [PubMed]
[38]. Xu B, Gottschalk W, Chow A, Wilson RI, Schnell E, Zang K, The role of brain-derived neurotrophic factor receptors in the mature hippocampus: modulation of long-term potentiation through a presynaptic mechanism involving TrkBJournal of Neuroscience 2000 20(18):6888-97.10.1523/JNEUROSCI.20-18-06888.200010995833 [Google Scholar] [CrossRef] [PubMed]
[39]. Hall J, Thomas KL, Everitt BJ, Rapid and selective induction of BDNF expression in the hippocampus during contextual learningNature Neuroscience 2000 3(6):533-35.10.1038/7569810816306 [Google Scholar] [CrossRef] [PubMed]
[40]. Saarelainen T, Lukkarinen JA, Koponen S, Gröhn OH, Jolkkonen J, Koponen E, Transgenic mice overexpressing truncated trkB neurotrophin receptors in neurons show increased susceptibility to cortical injury after focal cerebral ischemiaMolecular and Cellular Neuroscience 2000 16(2):87-96.10.1006/mcne.2000.086310924253 [Google Scholar] [CrossRef] [PubMed]
[41]. Egan MF, Kojima M, Callicott JH, Goldberg TE, Kolachana BS, Bertolino A, The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal functionCell 2003 112(2):257-69.10.1016/S0092-8674(03)00035-7 [Google Scholar] [CrossRef]
[42]. Pezet S, Malcangio M, Lever IJ, Perkinton MS, Thompson SW, Williams RJ, Noxious stimulation induces Trk receptor and downstream ERK phosphorylation in spinal dorsal hornMolecular and Cellular Neuroscience 2002 21(4):684-95.10.1006/mcne.2002.120512504600 [Google Scholar] [CrossRef] [PubMed]
[43]. Kerr B, Bradbury E, Bennett D, Trivedi P, Dassan P, French J, Brain-derived neurotrophic factor modulates nociceptive sensory inputs and NMDA-evoked responses in the rat spinal cordJournal of Neuroscience 1999 19(12):5138-48.10.1523/JNEUROSCI.19-12-05138.199910366647 [Google Scholar] [CrossRef] [PubMed]
[44]. Caporali A, Emanueli C, Cardiovascular actions of neurotrophinsPhysiological reviews 2009 89(1):279-308.10.1152/physrev.00007.200819126759 [Google Scholar] [CrossRef] [PubMed]
[45]. Meuchel LW, Thompson MA, Cassivi SD, Pabelick CM, Prakash Y, Neurotrophins induce nitric oxide generation in human pulmonary artery endothelial cellsCardiovascular research 2011 91(4):668-76.10.1093/cvr/cvr10721498417 [Google Scholar] [CrossRef] [PubMed]
[46]. Golden E, Emiliano A, Maudsley S, Windham BG, Carlson OD, Egan JM, Circulating brain-derived neurotrophic factor and indices of metabolic and cardiovascular health: data from the Baltimore Longitudinal Study of AgingPloS one 2010 5(4):e1009910.1371/journal.pone.001009920404913 [Google Scholar] [CrossRef] [PubMed]
[47]. Matthews V, Åström M-B, Chan M, Bruce C, Krabbe K, Prelovsek O, Brain-derived neurotrophic factor is produced by skeletal muscle cells in response to contraction and enhances fat oxidation via activation of AMP-activated protein kinaseDiabetologia 2009 52(7):1409-18.10.1007/s00125-009-1364-119387610 [Google Scholar] [CrossRef] [PubMed]
[48]. Steinman L, Elaborate interactions between the immune and nervous systemsNature Immunology 2004 5(6):575-81.10.1038/ni107815164017 [Google Scholar] [CrossRef] [PubMed]
[49]. Grande I, Fries GR, Kunz M, Kapczinski F, The role of BDNF as a mediator of neuroplasticity in bipolar disorderPsychiatry Investigation 2010 7(4):243-50.10.4306/pi.2010.7.4.24321253407 [Google Scholar] [CrossRef] [PubMed]
[50]. Jankowsky JL, Patterson PH, The role of cytokines and growth factors in seizures and their sequelaeProgress in Neurobiology 2001 63(2):125-49.10.1016/S0301-0082(00)00022-8 [Google Scholar] [CrossRef]
[51]. Lähteinen S, Pitkänen A, Saarelainen T, Nissinen J, Koponen E, Castrén E, Decreased BDNF signalling in transgenic mice reduces epileptogenesisEuropean Journal of Neuroscience 2002 15(4):721-34.10.1046/j.1460-9568.2002.01897.x11886452 [Google Scholar] [CrossRef] [PubMed]
[52]. Takahashi M, Hayashi S, Kakita A, Wakabayashi K, Fukuda M, Kameyama S, Patients with temporal lobe epilepsy show an increase in brain-derived neurotrophic factor protein and its correlation with neuropeptide YBrain Research 1999 818(2):579-82.10.1016/S0006-8993(98)01355-9 [Google Scholar] [CrossRef]
[53]. Binder DK, Croll SD, Gall CM, Scharfman HE, BDNF and epilepsy: too much of a good thing?Trends in neurosciences 2001 24(1):47-53.10.1016/S0166-2236(00)01682-9 [Google Scholar] [CrossRef]
[54]. Masliah E, Mallory M, Hansen L, Richard D, Alford M, Terry R, Synaptic and neuritic alterations during the progression of Alzheimer’s diseaseNeuroscience Letters 1994 174(1):67-72.10.1016/0304-3940(94)90121-X [Google Scholar] [CrossRef]
[55]. Laske C, Stransky E, Leyhe T, Eschweiler GW, Maetzler W, Wittorf A, BDNF serum and CSF concentrations in Alzheimer’s disease, normal pressure hydrocephalus and healthy controlsJournal of Psychiatric Research 2007 41(5):387-94.10.1016/j.jpsychires.2006.01.01416554070 [Google Scholar] [CrossRef] [PubMed]
[56]. Holsinger RD, Schnarr J, Henry P, Castelo VT, Fahnestock M, Quantitation of BDNF mRNA in human parietal cortex by competitive reverse transcription-polymerase chain reaction: decreased levels in Alzheimer’s diseaseMolecular Brain Research 2000 76(2):347-54.10.1016/S0169-328X(00)00023-1 [Google Scholar] [CrossRef]
[57]. Nagahara AH, Merrill DA, Coppola G, Tsukada S, Schroeder BE, Shaked GM, Neuroprotective effects of brain-derived neurotrophic factor in rodent and primate models of Alzheimer’s diseaseNature Medicine 2009 15(3):331-37.10.1038/nm.191219198615 [Google Scholar] [CrossRef] [PubMed]
[58]. Peng S, Garzon DJ, Marchese M, Klein W, Ginsberg SD, Francis BM, Decreased brain-derived neurotrophic factor depends on amyloid aggregation state in transgenic mouse models of Alzheimer’s diseaseJournal of Neuroscience 2009 29(29):9321-29.10.1523/JNEUROSCI.4736-08.200919625522 [Google Scholar] [CrossRef] [PubMed]
[59]. Blurton-Jones M, Kitazawa M, Martinez-Coria H, Castello NA, Müller F-J, Loring JF, Neural stem cells improve cognition via BDNF in a transgenic model of Alzheimer diseaseProceedings of the National Academy of Sciences 2009 106(32):13594-99.10.1073/pnas.090140210619633196 [Google Scholar] [CrossRef] [PubMed]
[60]. Reiner A, Albin RL, Anderson KD, D’Amato CJ, Penney JB, Young AB, Differential loss of striatal projection neurons in Huntington diseaseProceedings of the National Academy of Sciences 1988 85(15):5733-37.10.1073/pnas.85.15.57332456581 [Google Scholar] [CrossRef] [PubMed]
[61]. Zuccato C, Cattaneo E, Role of brain-derived neurotrophic factor in Huntington’s diseaseProgress in Neurobiology 2007 81(5):294-330.10.1016/j.pneurobio.2007.01.00317379385 [Google Scholar] [CrossRef] [PubMed]
[62]. Ferrer I, Goutan E, Marın C, Rey M, Ribalta T, Brain-derived neurotrophic factor in Huntington diseaseBrain Research 2000 866(1):257-61.10.1016/S0006-8993(00)02237-X [Google Scholar] [CrossRef]
[63]. Ciammola A, Sassone J, Cannella M, Calza S, Poletti B, Frati L, Low brain-derived neurotrophic factor (BDNF) levels in serum of Huntington’s disease patientsAmerican Journal of Medical Genetics Part B: Neuropsychiatric Genetics 2007 144(4):574-77.10.1002/ajmg.b.3050117427191 [Google Scholar] [CrossRef] [PubMed]
[64]. Spires TL, Grote HE, Varshney NK, Cordery PM, van Dellen A, Blakemore C, Environmental enrichment rescues protein deficits in a mouse model of Huntington’s disease, indicating a possible disease mechanismJournal of Neuroscience 2004 24(9):2270-76.10.1523/JNEUROSCI.1658-03.200414999077 [Google Scholar] [CrossRef] [PubMed]
[65]. Peng Q, Masuda N, Jiang M, Li Q, Zhao M, Ross CA, The antidepressant sertraline improves the phenotype, promotes neurogenesis and increases BDNF levels in the R6/2 Huntington’s disease mouse modelExperimental Neurology 2008 210(1):154-63.10.1016/j.expneurol.2007.10.01518096160 [Google Scholar] [CrossRef] [PubMed]
[66]. Shelton RC, The molecular neurobiology of depressionPsychiatric Clinics of North America 2007 30(1):1-11.10.1016/j.psc.2006.12.00517362799 [Google Scholar] [CrossRef] [PubMed]
[67]. NIH. Schizophrenia NIMH: NIH; 2016 [Google Scholar]
[68]. Vasic N, Connemann BJ, Wolf RC, Tumani H, Brettschneider J, Cerebrospinal fluid biomarker candidates of schizophrenia: where do we stand?European Archives of Psychiatry and Clinical Neuroscience 2012 262(5):375-91.10.1007/s00406-011-0280-922173848 [Google Scholar] [CrossRef] [PubMed]
[69]. Issa G, Wilson C, Terry AV, Pillai A, An inverse relationship between cortisol and BDNF levels in schizophrenia: data from human postmortem and animal studiesNeurobiology of Disease 2010 39(3):327-33.10.1016/j.nbd.2010.04.01720451611 [Google Scholar] [CrossRef] [PubMed]
[70]. Pillai A, Terry AV, Mahadik SP, Differential effects of long-term treatment with typical and atypical antipsychotics on NGF and BDNF levels in rat striatum and hippocampusSchizophrenia Research 2006 82(1):95-106.10.1016/j.schres.2005.11.02116442781 [Google Scholar] [CrossRef] [PubMed]
[71]. Knable MB, Barci BM, Webster MJ, Meador-Woodruff J, Torrey EF, Molecular abnormalities of the hippocampus in severe psychiatric illness: postmortem findings from the Stanley Neuropathology ConsortiumMolecular Psychiatry 2004 9(6):609-20.10.1038/sj.mp.400147114708030 [Google Scholar] [CrossRef] [PubMed]
[72]. Hovatta I, Barlow C, Molecular genetics of anxiety in mice and menAnnals of Medicine 2008 40(2):92-109.10.1080/0785389070174709618293140 [Google Scholar] [CrossRef] [PubMed]
[73]. Monteggia LM, Luikart B, Barrot M, Theobold D, Malkovska I, Nef S, Brain-derived neurotrophic factor conditional knockouts show gender differences in depression-related behavioursBiological Psychiatry 2007 61(2):187-97.10.1016/j.biopsych.2006.03.02116697351 [Google Scholar] [CrossRef] [PubMed]
[74]. Autry AE, Adachi M, Cheng P, Monteggia LM, Gender-specific impact of brain-derived neurotrophic factor signaling on stress-induced depression-like behaviourBiological psychiatry 2009 66(1):84-90.10.1016/j.biopsych.2009.02.00719358977 [Google Scholar] [CrossRef] [PubMed]
[75]. Chen Z-Y, Jing D, Bath KG, Ieraci A, Khan T, Siao C-J, Genetic variant BDNF (Val66Met) polymorphism alters anxiety-related behaviourScience 2006 314(5796):140-43.10.1126/science.112966317023662 [Google Scholar] [CrossRef] [PubMed]
[76]. Frustaci A, Pozzi G, Gianfagna F, Manzoli L, Boccia S, Meta-analysis of the brain-derived neurotrophic factor gene (BDNF) Val66Met polymorphism in anxiety disorders and anxiety-related personality traitsNeuropsychobiology 2008 58(3-4):163-70.10.1159/00018289219088493 [Google Scholar] [CrossRef] [PubMed]
[77]. Soliman F, Glatt CE, Bath KG, Levita L, Jones RM, Pattwell SS, A genetic variant BDNF polymorphism alters extinction learning in both mouse and humanScience 2010 327(5967):863-66.10.1126/science.118188620075215 [Google Scholar] [CrossRef] [PubMed]
[78]. Frielingsdorf H, Bath KG, Soliman F, DiFede J, Casey B, Lee FS, Variant brain-derived neurotrophic factor Val66Met endophenotypes: implications for posttraumatic stress disorderAnnals of the New York Academy of Sciences 2010 1208(1):150-57.10.1111/j.1749-6632.2010.05722.x20955337 [Google Scholar] [CrossRef] [PubMed]
[79]. Monteggia LM, Barrot M, Powell CM, Berton O, Galanis V, Gemelli T, Essential role of brain-derived neurotrophic factor in adult hippocampal functionProceedings of the National Academy of Sciences of the United States of America 2004 101(29):10827-32.10.1073/pnas.040214110115249684 [Google Scholar] [CrossRef] [PubMed]
[80]. Rattiner LM, Davis M, French CT, Ressler KJ, Brain-derived neurotrophic factor and tyrosine kinase receptor B involvement in amygdala-dependent fear conditioningJournal of Neuroscience 2004 24(20):4796-806.10.1523/JNEUROSCI.5654-03.200415152040 [Google Scholar] [CrossRef] [PubMed]
[81]. Heldt S, Stanek L, Chhatwal J, Ressler K, Hippocampus-specific deletion of BDNF in adult mice impairs spatial memory and extinction of aversive memoriesMolecular Psychiatry 2007 12(7):656-70.10.1038/sj.mp.400195717264839 [Google Scholar] [CrossRef] [PubMed]
[82]. Smith MA, Makino S, Kim S, Kvetnansky R, Stress increases brain-derived neurotropic factor messenger ribonucleic acid in the hypothalamus and pituitaryEndocrinology 1995 136(9):3743-50.10.1210/endo.136.9.76490807649080 [Google Scholar] [CrossRef] [PubMed]
[83]. Rios M, Fan G, Fekete C, Kelly J, Bates B, Kuehn R, Conditional deletion of brain-derived neurotrophic factor in the postnatal brain leads to obesity and hyperactivityMolecular endocrinology 2001 15(10):1748-57.10.1210/mend.15.10.070611579207 [Google Scholar] [CrossRef] [PubMed]
[84]. Das UN, Obesity: genes, brain, gut, and environmentNutrition 2010 26(5):459-73.10.1016/j.nut.2009.09.02020022465 [Google Scholar] [CrossRef] [PubMed]
[85]. Tsuchida A, Nonomura T, Ono-Kishino M, Nakagawa T, Taiji M, Noguchi H, Acute effects of brain-derived neurotrophic factor on energy expenditure in obese diabetic miceInternational Journal of Obesity & Related Metabolic Disorders 2001 25(9)10.1038/sj.ijo.080167811571589 [Google Scholar] [CrossRef] [PubMed]
[86]. Tonra JR, Ono M, Liu X, Garcia K, Jackson C, Yancopoulos GD, Brain-derived neurotrophic factor improves blood glucose control and alleviates fasting hyperglycaemia in C57BLKS-Lepr (db)/lepr (db) miceDiabetes 1999 48(3):588-94.10.2337/diabetes.48.3.58810078561 [Google Scholar] [CrossRef] [PubMed]
[87]. Ono M, Itakura Y, Nonomura T, Nakagawa T, Nakayama C, Taiji M, Intermittent administration of brain-derived neurotrophic factor ameliorates glucose metabolism in obese diabetic miceMetabolism 2000 49(1):129-33.10.1016/S0026-0495(00)90988-0 [Google Scholar] [CrossRef]
[88]. Jonas JB, Bourne RR, White RA, Flaxman SR, Keeffe J, Leasher J, Visual impairment and blindness due to macular diseases globally: a systematic review and meta-analysisAmerican Journal of Ophthalmology 2014 158(4):808-15.10.1016/j.ajo.2014.06.01224973605 [Google Scholar] [CrossRef] [PubMed]
[89]. Johnson JE, Barde Y-A, Schwab M, Thoenen H, Brain-derived neurotrophic factor supports the survival of cultured rat retinal ganglion cellsJournal of Neuroscience 1986 6(10):3031-38.10.1523/JNEUROSCI.06-10-03031.19862876066 [Google Scholar] [CrossRef] [PubMed]
[90]. Cellerino A, Kohler K, Brain-derived neurotrophic factor/neurotrophin-4 receptor TrkB is localized on ganglion cells and dopaminergic amacrine cells in the vertebrate retinaJournal of Comparative Neurology 1997 386(1):149-60.10.1002/(SICI)1096-9861(19970915)386:1<149::AID-CNE13>3.0.CO;2-F [Google Scholar] [CrossRef]
[91]. Khalin I, Alyautdin R, Kocherga G, Bakar MA, Targeted delivery of brain-derived neurotrophic factor for the treatment of blindness and deafnessInternational journal of nanomedicine 2015 10:324510.2147/IJN.S7748025995632 [Google Scholar] [CrossRef] [PubMed]
[92]. Zuccato C, Cattaneo E, Brain-derived neurotrophic factor in neurodegenerative diseasesNature Reviews Neurology 2009 5(6):31110.1038/nrneurol.2009.5419498435 [Google Scholar] [CrossRef] [PubMed]
[93]. Chen K-W, Chen L, Epigenetic regulation of BDNF gene during development and diseasesInternational journal of Molecular Sciences 2017 18(3):57110.3390/ijms1803057128272318 [Google Scholar] [CrossRef] [PubMed]