The extensor apparatus of knee is composed of Quadriceps Femoris (QF) muscle, patella and ligamentum patellae.
The QF is great extensor muscle of the leg which is divided into four parts; one is rectus femoris and other three vasti, VM, VL and Vastus Intermedius (VI). All four components of QF unite in the lower part of the thigh to form a single strong tendon attached to the patella and some fibers continue over it to blend with the patellar tendon. VM arises from the lower part of the intertrochanteric line, spiral line, medial lip of linea aspera, proximal part of medial supracondylar line, the adductor longus, adductor magnus muscles and medial intermuscular septum. Its fibers pass downwards and forwards and are attached to the QF tendon and medial border of patella. The lowest fibers of VM are nearly horizontal. Some authors distinguish this part of muscle as VMO that originates from the tendon of adductor magnus and insert into the medial border of patella, it plays an important role in the function of patello femoral joint [1].
The quadriceps force vector includes forces from the fiber orientation of VL, VI, Rectus Femoris (RF) and VM. The VL is composed of two force vector components, the VLL and VLO. The VM is composed of two force vector components, the VML and VMO. In the coronal plane, the QF force angles are made by the VLO at 35 degrees and VLL at 14 degrees laterally, by the VI and RF at 0 degree and by VMO at 47 degrees and VML at 15 degrees medially [2]. Hence, extremely important dynamic function of VMO is to counter this lateral vector on the patella during knee extension.
Proper alignment and restoration of the kinematic variable in the patellofemoral mechanism are essential for adequate functioning and longevity of joint [3]. Patelofemoral pain is common in general population, more common in women and athletes [4]. Atrophy of VMO has often been suggested to play an important role in pathophysiology of unstable patello femoral joint [5]. The VM is the key to the knee because it is almost entirely responsible for stabilization and protection. Non operative treatment of Patello Femoral Pain Syndrome (PFPS) and many rehabilitation programmes focus primarily on strengthening the VMO [6]. So, VMO presents a biomechanical advantage over the VL due to oblique orientation of its muscle fibers.
For total knee replacement several approaches have been aimed to position the arthrotomy incision in such a way that the patella can be retracted without violation of the quadriceps tendon or of vastus medialis [7]. This approach is less damaging to VM, it results in less blood loss and fast recovery [8]. Hence, structural parameters of the VM are useful for both surgical and conservative treatment of knee pathology.
The aim of our study was to determine muscle fiber angles and lengths of VML and VMO, to find out the distinct nerve supply and fascial plane between VML and VMO and to evaluate VM insertion in relation to patella.
Materials and Methods
The present prospective observational study was carried out on 30 intact lower limbs (right-15 and left-15) from 15 donated cadavers, (13 males and 2 females) with an age 60 years and above in the Department of Anatomy, Medical College Baroda, Gujarat, India, during July 2016 to March 2017. The cadavers had no visible evidence of surgery or trauma to the lower limbs and also no disruption of muscular anatomy at hip or knee.
Each cadaver was placed in supine position with the patella upwards. The dissection method was obtained from previous study [12]. Two horizontal skin incisions were made, one, upper at the level of Anterior Superior Iliac Spine (ASIS) and another at the level of Tibial Tuberosity (TT). One vertical incision was made from ASIS to TT; blunt dissection was done to separate the underlying fascia. Sartorius muscle was separated from deeper structures and four parts of quadriceps muscle and tendon were separated from each other. Sartorius muscle was cut at the level of intersection point with the VM muscle and retracted medially.
Four marker pins were inserted in the VM as below [Table/Fig-1a]:
Measurement methods of vastus medialis muscle; a) Showing parts of VM muscle; b) Measurement of VM length with flexible ruler; c) Measurement of VM angle with goniometer.
P= Anterior superior iliac spine, Q= Tibial tuberosity, VM= Vastus medialis muscle, RF= Rectus femoris muscle, White thread indicates femoral axis
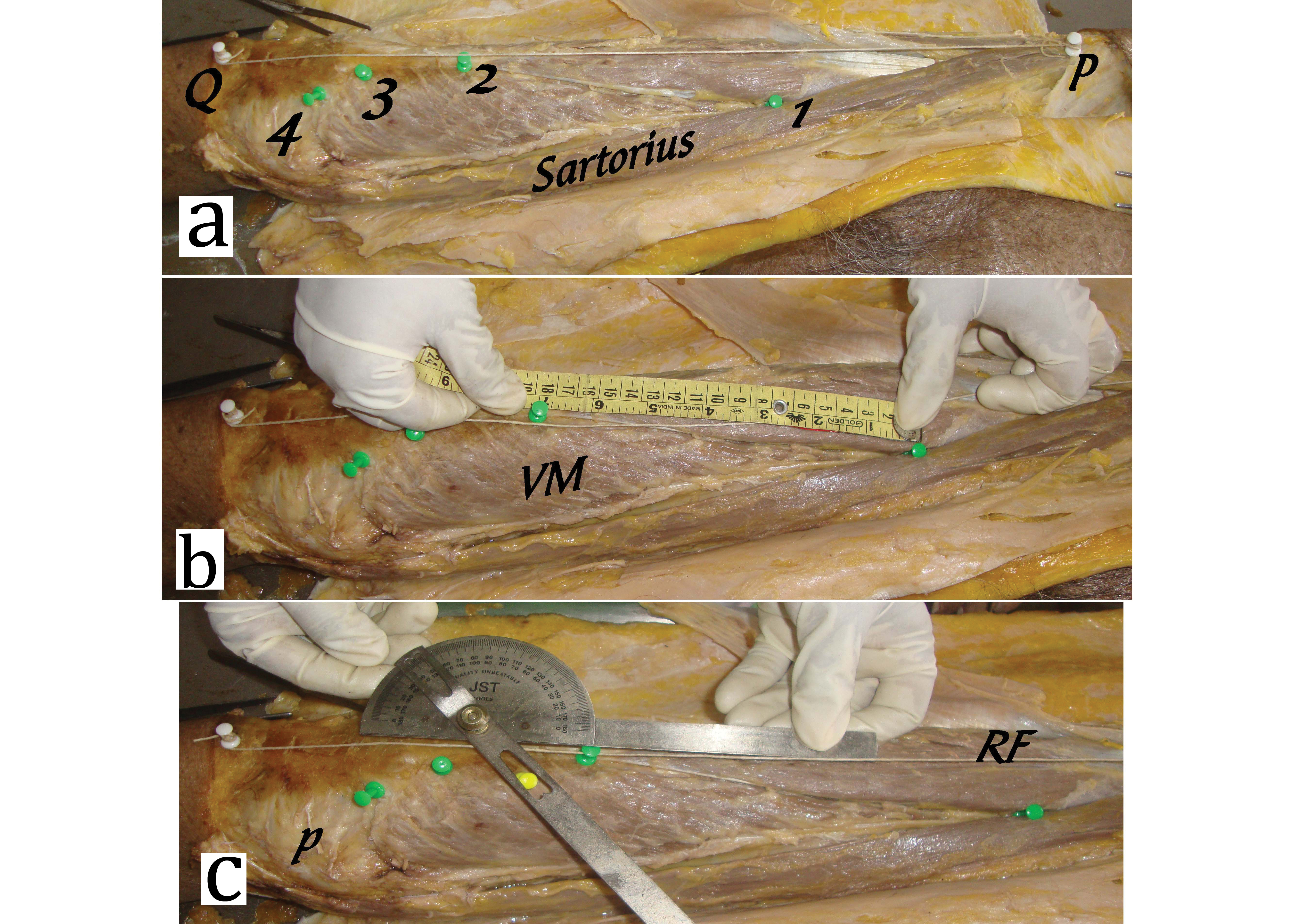
First at the intersection point of VM with the sartorius muscle;
Second at the upper most point of muscle fibers inserted into the quadriceps tendon;
Third at the muscle fiber insertion at the superior aspect of patella;
Last at the lower most point of muscle fiber insertion on the medial aspect of patella.
These marker pins divides VM muscle visually into three parts. Part between first and second pins is VML. Part between second and fourth pins is VMO. VMO is further divided into VMO superior (VMOs) between second and third pins and VMO inferior (VMOi) between third and fourth pins. Lengths of each part were measured with the help of flexible ruler in centimetres [Table/Fig-1b].
Two additional pins were inserted, first at the ASIS and second at the midpoint of TT. The thread was tied between these two points. This determines femoral axis. Angle of muscle fibers measured with the help of Goniometer at the midpoint of each part of VM muscle [Table/Fig-1c].
The fascial plane and the femoral nerve and its branches supplying VM were dissected and terminal branches were traced in each section with the blunt dissection.
Lengths of VML, VMOs and VMOi were calculated as a percentage of total VM length. Fiber angle for each part of VM was determined accordingly to the femoral axis. Nerve planes and fascial planes were recorded.
Statistical Analysis
Descriptive statistics were calculated with the help of statistical expert for all of the measurements and statistical differences were analysed by using a one-way ANOVA MedCalc version 12.5. The level of significance was set at a p<0.05.
Results
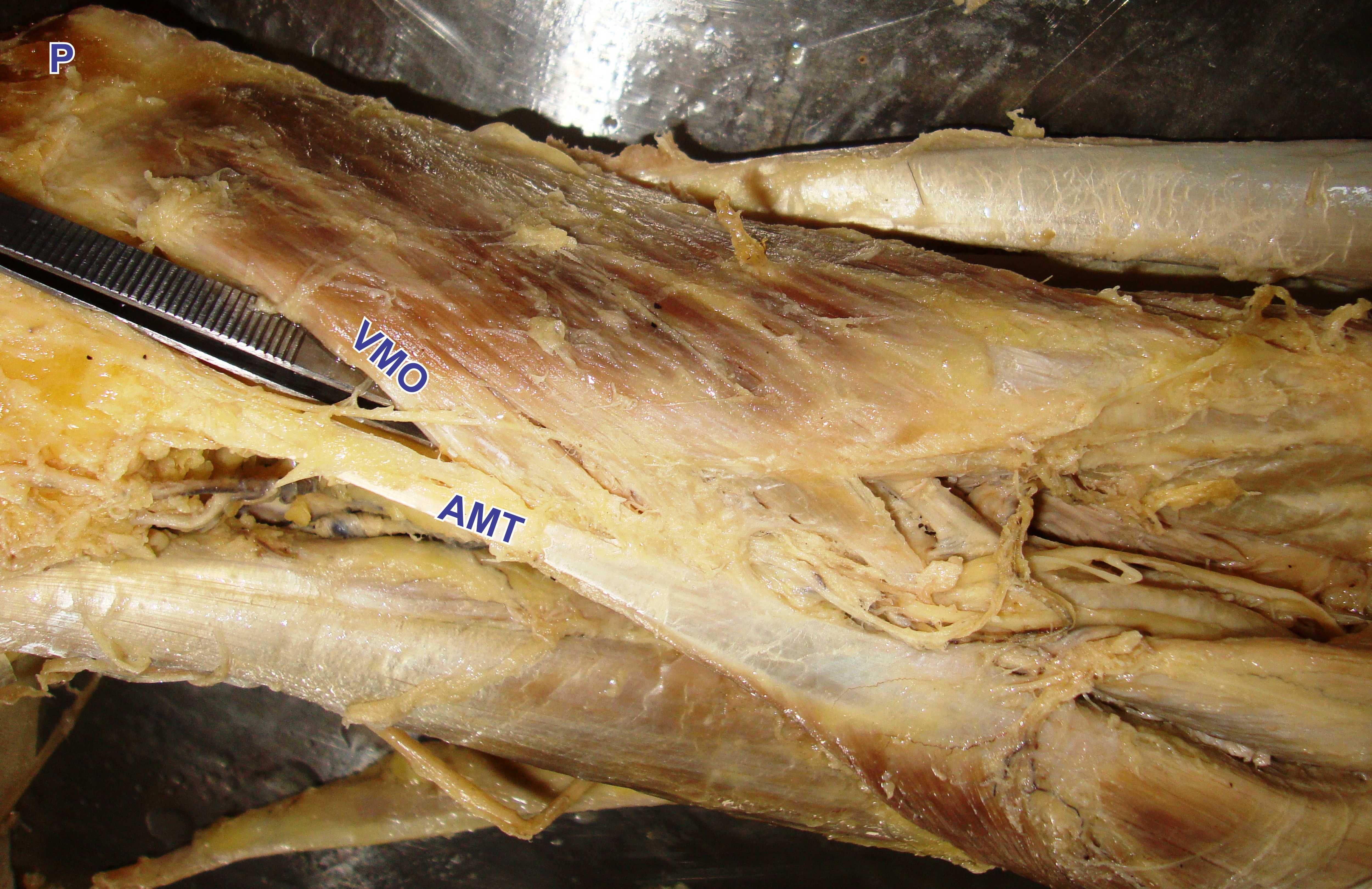
In all the limbs, the VM muscle consists of long proximal portion (VML), and an oblique distal portion (VMO). The VML fibers join the quadriceps tendon around 8-12 cm above the superior border of patella. The VMO fibers took origin from the adductor magnus muscle and fibers attached to the patella [Table/Fig-2]. These fibers further divided into VMOs fibers, attached to the superior border of the patella while VMOi fibers were attached to medial border of patella. Insertion of VMO in relation to medial border of patella was observed. In 16.66% cases insertion was extended up to upper one third, in 63.66% cases upper two third while in 3.33% cases whole medial border of patella. In rest 16.66% cases VMO inserted only to upper border of patella and these cases labelled as absent VMOi. Hence, VMOs present in all the cases while VMOi was present in 25 limbs (83.33%) and absent in five limbs (16.66%).
Origin of VMO from adductor magnus tendon.
VMO= Vastus medialis obliquus, AMT= Adductor magnus tendon, P= Patella
Compared to total VM length, the range of VML, VMOs and VMOi lengths are 14.5-29 cm, 5-10 cm and 1.5-5 cm respectively. The fiber angles ranged for VML, VMOs and VMOi were between 18 to 32, 30 to 46 and 46 to 56 degrees respectively. The mean value and standard deviations for fiber angles and section length in relation with total VM length were calculated. There were significant differences among all groups (p=0.001) [Table/Fig-3].
Vastus medialis fiber angle and section percentages of total VM length.
| Variables | Fiber Angles* [degree]Mean ±SD | Section length/Total length [%]Mean ±SD |
|---|
| VML | 24.16 ± 4.71 | 65.54 ± 4.84 |
| VMO superior | 35.8 ±5.09 | 25.23 ± 4.20 |
| VMO inferior | 41.26± 3.67 | 9.22 ± 2.17 |
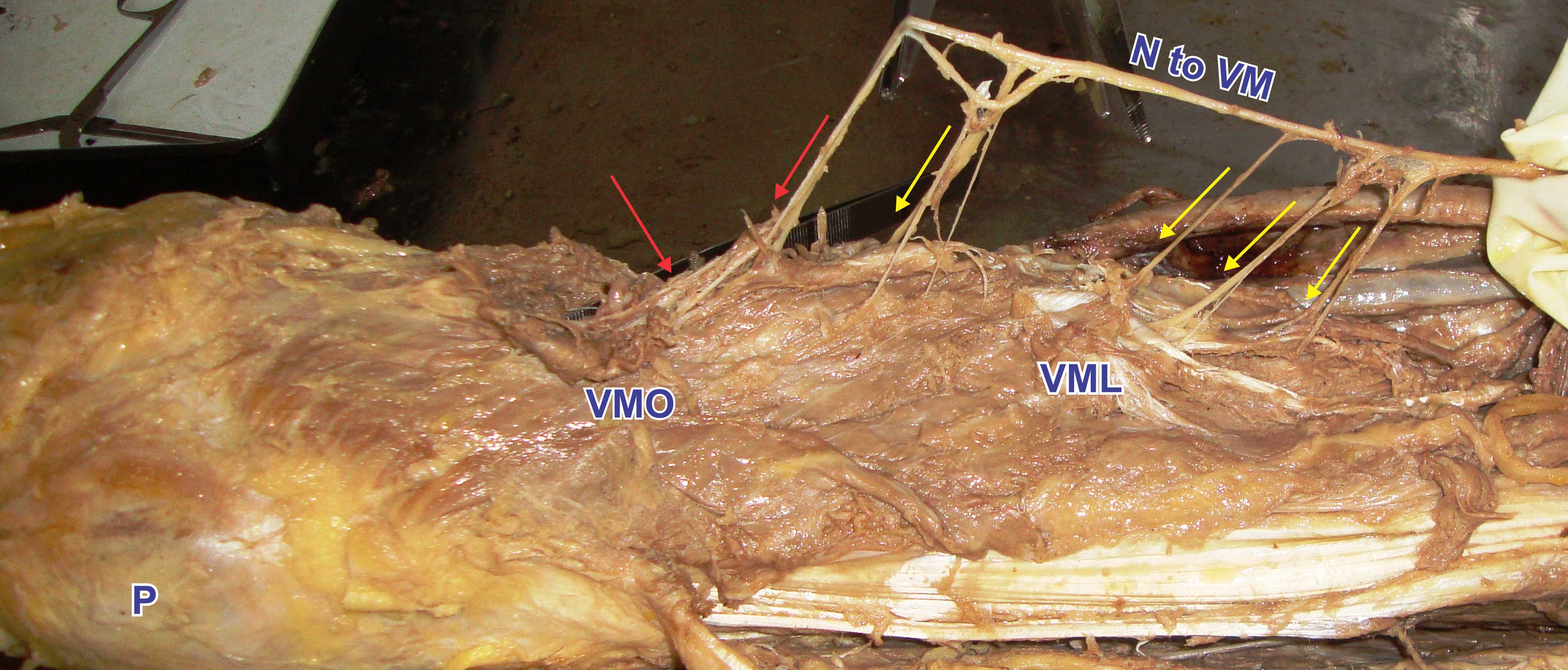
In all the specimens, nerve to VM, a branch of posterior division of femoral nerve run in a deeper aspect of muscle, showed typical pattern of distribution. The nerve divides into short lateral and long medial branches. The lateral branch divides into 2 to 3 branches and supplies upper part of VM while long medial branch runs distally along the anteromedial border of muscle. In its course it gives 3 to 4 slender branches to middle part of muscle and in distal part it is sprayed into 4 to 5 terminal branches [Table/Fig-4]. This shows that the distal part of VM receives more numbers of terminal branches as compared to proximal part. There was no distinct fascial plane found between VML and VMO.
Nerve supply of VM muscle.
N to VM= Nerve to vastus medialis muscle, yellow arrows indicate branches from VML, red arrows indicate branches from VMO.
In present study, the data for right and left sides were also compared. On right side, the range of VML, VMOs and VMOi lengths were 15-29 cm, 5-10 cm and 1.5-5 cm while on left side lengths were 14.5-28 cm, 6-10 cm and 1.5-4 cm respectively. The fiber angles ranged for VML, VMOs, VMOi were 18-32, 30-46 and 46-56 degrees on right side while on left side it was 18-32, 30-46 and 46-54 degree respectively. A p-value for the length was 0.006 while for angles it was 0.009 which were found significant.
The present study cannot comment on gender differences of VM muscle, as this study includes only two female cadavers.
Though, the fiber angles progressively increased from VML to VMO but there was not any abrupt change in fiber direction. Also, there was no separate nerve supply and fascial plane separating VML and VMO.
Discussion
The present study confirms and clarifies past studies and has demonstrated that the VM should be classified anatomically and functionally as a single muscle, although there is accurate description found in various literatures that the VM consists of two parts, long and oblique parts based on the variation in the angle of muscle fibre orientation [9,12,13]. The clinical theory describes that the two parts of VM muscles have different functions, but there were very few anatomical evidence to support this.
Carlson K et al., found the mean VML, VMOs and VMOi fiber angles, respectively, are 210±4, 300±3 and 570±4 [12]. In the present study proximal long part of VM (VML) had more vertically oriented fibres (angle range-18-32 degrees) while distal oblique part of VM (VMO) with more horizontally oriented fibres (angle range 30-56 degree). This shows fiber angles were increasing downwards. Hubbard JK et al., found that the average fiber angle of VML was 6-28 degrees and of VMO was 28-70 degrees [14]. This range difference in Hubbard study is may be due to large sample size. There was not an abrupt change in fiber orientation which would demarcate VML and VMO as separate muscles.
In all the cases present study showed the distal part of VM muscle originated from adductor magnus tendon. The origin of distal part of the VM from the adductor magnus was a human adaptation to bipedal gait, unique to human and absent in other primates [14].
There were variable extents of insertion of VM on medial border of patella. Chavan SK et al., found that in 80% cases insertion extended to the upper two third in 60% or whole (20%) of medial margin of patella. While in another 20% of cases the extent is higher than what has been described [15]. In the present study, in 63.66 % cases insertion extended up to upper two third while in 3.33% cases whole medial border of patella.
In 16.66% cases the insertion extended up to upper one third and in rest of 16.66% cases insertion was limited to superior border of patella. No data was found in literature regarding absent VMOi. If such cases of absent VMOi correlated functionally, these cases may be more susceptible to anterior knee pain due to inability to counteract the normal lateral traction exerted by quadriceps tendon [15].
There were several studies reporting different findings regarding the presence of separate fascial plane and innervations that would distinguish the long and oblique heads of the VM as separate muscles which allow independent function [13,15,16]. But this was present in only few percentages of cadaver dissections. Hubbard JK et al., also found a distinct fascial plane [14]. Bose K et al., did not find any separating fascial plane [11]. Gunal I et al., found that in all 60 cadavers VMO receive additional nerve supply from saphenous nerve [17]. Other researchers found that VMO was only supplied by a branch of femoral nerve [12,13,17]. Study by Rajani T found the lowermost muscle fibers receiving the richest nerve supply [16].
The present study also confirmed that VM receives branches only from femoral nerve and no additional supply from saphenous nerve. The richest nerve supply to VMO may explain a special functional role of VMO in extension and stabilization of knee. There was no distinct fascial plane separating proximal and distal part of VM. Observations of present study were compared with past cadaveric studies given in [Table/Fig-5] [12,14,15,18,19].
Comparison of present study with the past studies.
| Authors | Nozic M et al., (1997) [18] | Hubbard JK et al., (1997) [14] | Peeler J et al., (2005) [19] | Carlson K et al., (2012) [12] | Chavan SK et al., (2016) [15] | Present study (2017) |
|---|
| No of Limbs | 50 | 374 | 32 | 02 | 40 | 30 |
| VML angle | 110±3 | 140±4 | 220±5 | 210±4 | - | 24.160±4.71 |
| VMO superior angle | - | - | 370±6 | 300±3 | - | 35.80±5.09 |
| VMO inferior angle | 520±6 | 530±7 | 570±6 | 570±4 | - | 41.260±3.67 |
| Fascial plane | 1/50 | 81/374 | 0/32 | 0/2 | 15/40 | 0/30 |
| Innervations | 1/50 | - | 0/32 | 0/2 | 23/40 | 0/30 |
The present study showed the significant difference on right and left sides. In contrast, previous studies did not find any statistical difference in gender and laterality [14,20,21]. This could be due to variations in preferential use of the lower limb which is influenced by genetical and environmental factors [22]. In the present study we did not have details of lifestyle, handedness and footedness and sports habits of the cadavers during their life.
Limitation
Limitation of present study was gender differences cannot be calculated due to less numbers of female cadavers. This study can be correlated in future with age related changes in VMO insertion and functional aspect of VM muscle in the cases of absent VMO inferior.
Conclusion
The present study concludes that the VM is having two parts VML and VMO based on the variation in the angle of muscle fiber orientation. These two parts were not separated by distinct fascial plane and no separate nerve supply to label it anatomically separate entity. But VMO has obtuse angled, horizontal fibers and richest nerve supply as compared to VML, also has special role in medial stabilization of patella. These data will be useful to the surgeons when there are obvious benefits of preserving the VMO. Present study also noted absent VMOi in 16.66% cases which is a unique finding. Such cases need to be correlated clinically and functionally.